Sunlight provides the energy for photosynthesis
Whether in water or on land, energy from the Sun is essential for the existence of most life on Earth. To understand how this energy is captured, we need to examine the energy that is available, the energy that is absorbed, and how this energy is converted into a usable form by photosynthesis. Plants have evolved several adaptive approaches for conducting photosynthesis in terrestrial environments; these mechanisms coincide with the environmental conditions of different regions of the world.
Available and Absorbed Solar Energy
Electromagnetic radiation Energy from the Sun, packaged in small particle-like units called photons.
The energy from the Sun, known as electromagnetic radiation, is packaged in small particle-like units called photons. The energy of photons is positively related to their frequency and inversely to their wavelength; the highest energy photons have the highest frequency and the shortest wavelengths. Wavelengths are expressed in units of nanometers (nm); a nanometer is one-billionth of a meter. The different wavelengths of light can be separated using a prism. As you can see in Figure 3.7, infrared radiation has long wavelengths, which we know contain lower energy. Short wavelengths, such as ultraviolet radiation, contain higher energy. Between the two extremes of infrared and ultraviolet radiation are wavelengths collectively known as visible light, which are visible to the human eye. Visible light represents only a small part of the spectrum of electromagnetic radiation.
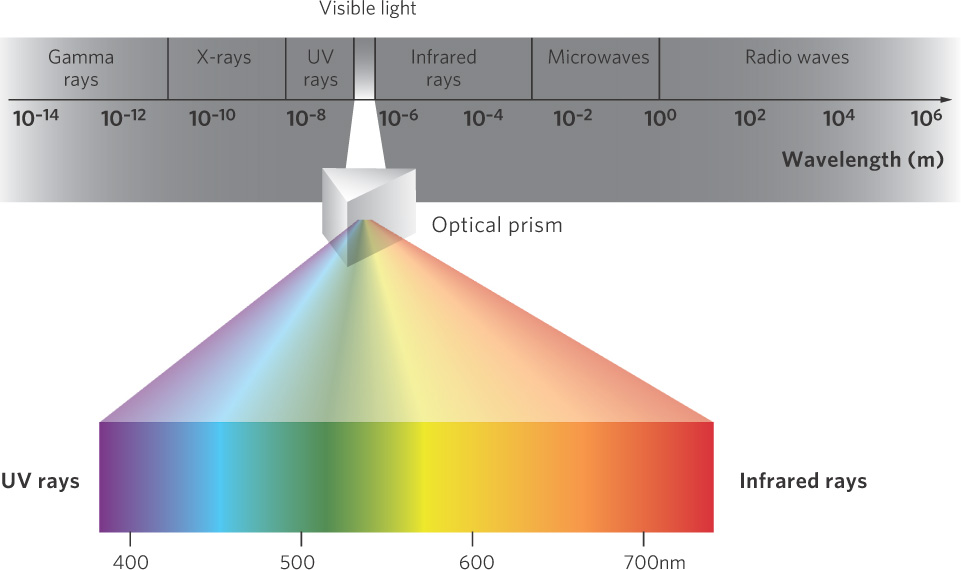
Visible light Wavelengths in between infrared and ultraviolet radiation that are visible to the human eye.
65
Photosynthetically active region Wavelengths of light that are suitable for photosynthesis.
The visible portion of the spectrum includes the photosynthetically active region, which consists of wavelengths of light that are suitable for photosynthesis. This range of wavelengths falls between about 400 nm (violet) and 700 nm (red). Plants, algae, and some bacteria absorb these wavelengths and assimilate their energy by photosynthesis. These also happen to be the wavelengths of greatest intensity at the Earth’s surface.
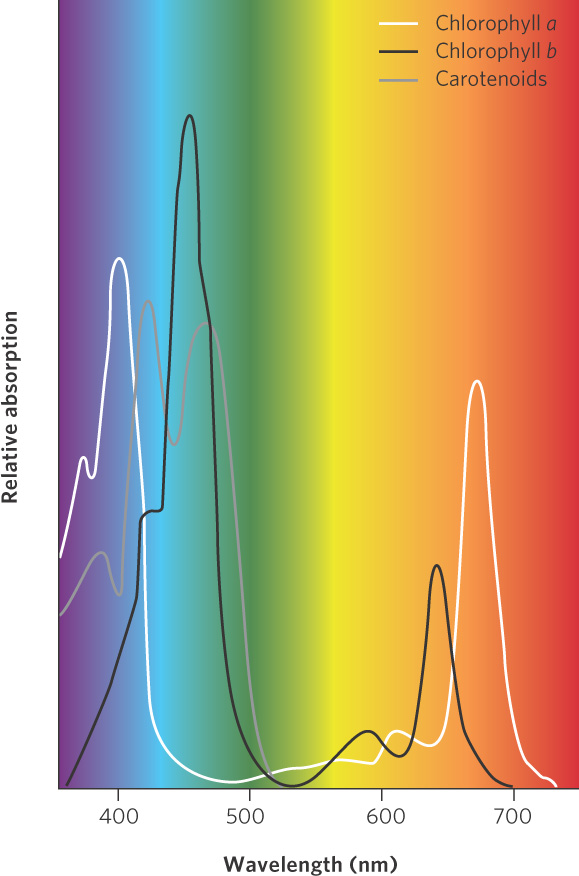
Eukaryotic photosynthetic organisms contain specialized cell organelles known as chloroplasts. As you can see in Figure 3.8, chloroplasts contain stacks of membranes known as thylakoids and a fluid-filled space surrounding the thylakoids called the stroma. Embedded within the thylakoid membranes are several kinds of pigments that absorb solar radiation, including chlorophylls and carotenoids. The patterns of absorption for several of these pigments are shown in Figure 3.9.
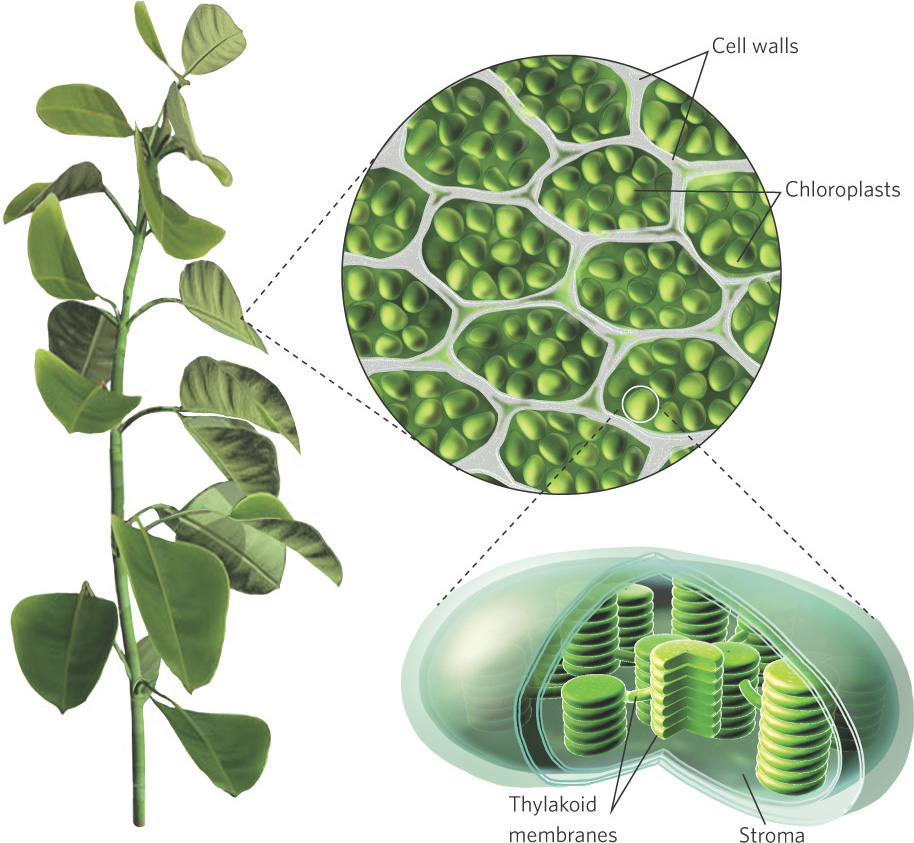
Chloroplasts Specialized cell organelles found in photosynthetic organisms.
Chlorophylls, which are primarily responsible for capturing light energy for photosynthesis, absorb red and violet light. Chlorophylls reflect green and blue light, which is why leaves on most plants are predominantly green in color. Over the past 60 years, scientists identified four types of chlorophyll that differ in the wavelengths that they absorb: chlorophyll a, b, c, and d. Chlorophyll a is found in all photosynthesizing organisms and is responsible for the actual steps in photosynthesis. The other types of chlorophyll act as accessory pigments, meaning that they capture light energy and then pass it on to chlorophyll a. In 2010, however, scientists reported the discovery of a fifth type of chlorophyll that they named chlorophyll f. This pigment, discovered in algae that live in shallow rocky pools on the coast of Australia, absorbs light at longer wavelengths than the others.
66
Carotenoids are also accessory pigments and they include two major classes: carotenes and xanthophylls. Carotenoids absorb primarily blue and green light, thereby complementing the absorption spectrum of chlorophyll. Carotenoids, the pigments that give carrots their orange color, reflect yellow and orange light. By containing carotenoids and several types of chlorophyll, producers can absorb a wider range of solar energy and use it to power photosynthesis.
Photosynthesis
In this section, we review the process of photosynthesis and the different pathways that have evolved under different ecological conditions. During photosynthesis, photosynthetic pigments absorb energy from photons of light, which is then converted into chemical energy stored in the high-energy bonds of organic compounds. In its simplest form, photosynthesis is the process of combining CO2, H2O, and solar energy to produce glucose (C6H12O6) and oxygen:
6 CO2 + 6 H2O + photons → C6H12O6 + 6 02
This simple equation summarizes a long chain of complex chemical reactions. As shown in Figure 3.10, the process takes place in two parts: light reactions and the Calvin cycle.
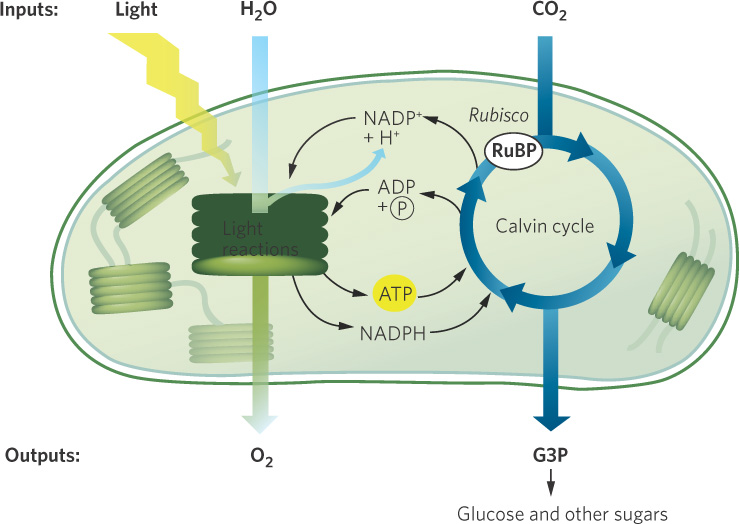
Light reactions depend on light energy from the Sun and include a series of events from the absorption of light to the production of high-energy compounds and oxygen (O2). First, photosynthetic pigments inside the thylakoids capture light energy. When chlorophyll molecules absorb photons, they release electrons, which are then passed along a chain of reactions. In the process, H2O molecules are split into hydrogen (H+) and oxygen (O2−) atoms. The individual oxygen atoms immediately combine with each other to become molecular oxygen (O2). The energy absorbed from the released electrons, as well as the H+ ions that are split from H2O, are used to produce two high-energy compounds. ADP (adenosine diphosphate) and P (phosphate) combine to form ATP (adenosine triphosphate), whereas NADP+ (nicotinamide adenine dinucleotide phosphate) and H+ combine to form NADPH (the reduced form of NADP+).
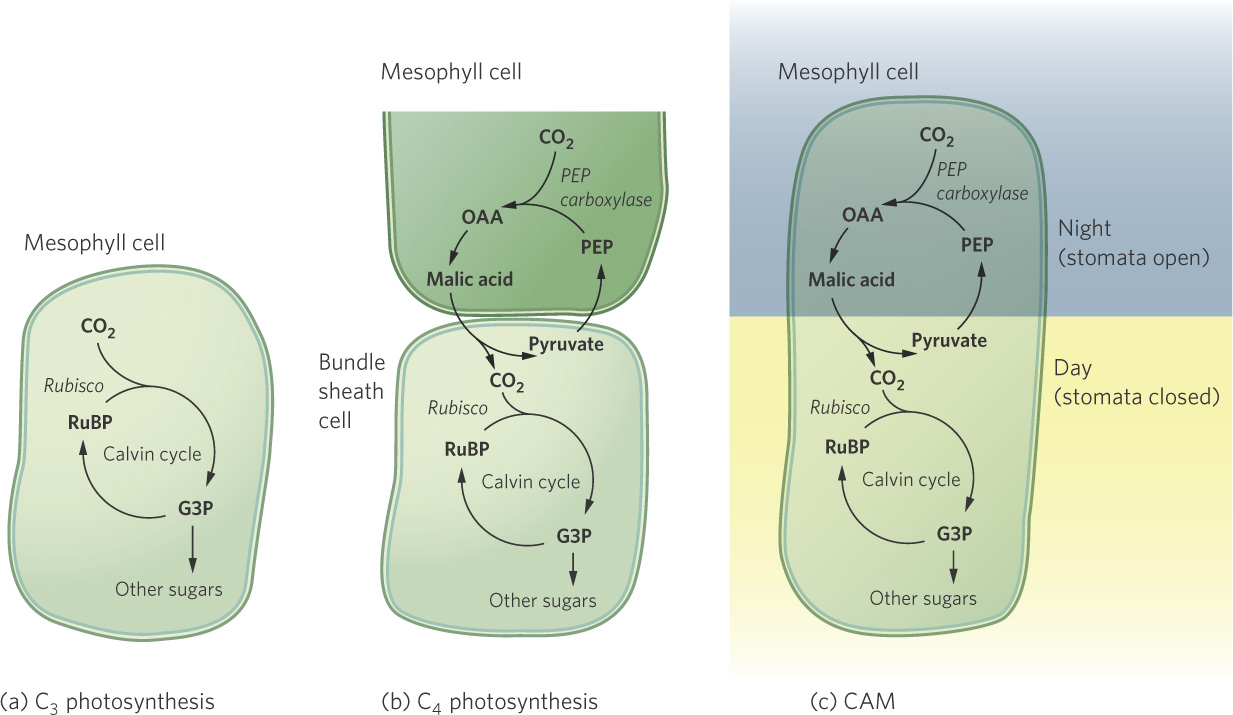
Once the cell has created the high-energy compounds ATP and NADPH, it uses the energy in these compounds to convert CO2 into glucose in a process known as the Calvin cycle. This takes place in the stroma of the chloroplast. Over evolutionary time, three distinct biochemical pathways have evolved for the Calvin cycle: C3, C4, and CAM photosynthesis. As we will soon see, each of these pathways is suited to particular ecological conditions.
C₃Photosynthesis
For most plants, photosynthesis begins with a reaction between CO2 and a five-carbon sugar known as RuBP (ribulose bisphosphate) to produce a six-carbon carbon compound. This reaction is catalyzed by the enzyme RuBP carboxylase-oxidase (also known as Rubisco). Once the six-carbon compound is created, it immediately splits into two molecules of a three-carbon sugar called G3P (glyceraldehyde 3-phosphate). We can represent this process as:
RuBP carboxylase-oxidase An enzyme involved in photosynthesis that catalyzes the reaction of RuBP and CO2 to form two molecules of glyceraldehyde 3-phosphate (G3P). Also known as Rubisco.
CO2 + RuBP → 2 G3P
C3 photosynthesis The most common photosynthetic pathway, in which CO2 is initially assimilated into a three-carbon compound, glyceraldehyde 3-phosphate (G3P).
This photosynthetic pathway, in which CO2 is initially assimilated into the three-carbon compound (G3P), is known as C3 photosynthesis. The vast majority of plants on Earth use C3 photosynthesis. In most plants, this process occurs in mesophyll cells in leaves.
67
One of the challenges for plants using C3 photosynthesis is that Rubisco, the enzyme responsible for joining CO2 and RuBP, has a low affinity for CO2 compared to another enzyme that we will discuss shortly. Consequently, carbon assimilation using Rubisco is quite inefficient at the low concentrations of CO2 found in the mesophyll cells of plant leaves. To achieve high rates of carbon assimilation, plants must therefore pack their mesophyll cells with large amounts of the enzyme Rubisco. In some plant species, Rubisco can constitute up to 30 percent of the dry weight of leaf tissue.
Rubisco’s low affinity for CO2 is not the only problem plants face. Under certain conditions—such as when temperatures are high, concentrations of O2 are high, and CO2 concentrations are low—Rubisco preferentially binds to O2 rather than CO2. This occurs when hot, dry conditions cause the leaf stomata to close, preventing new CO2 from entering to replenish the CO2 that has been consumed by the Calvin cycle. Closed stomata also prevent O2 produced by the light reaction from leaving the leaf. Consequently, hot and dry conditions lead to changes in CO2 and O2 concentrations that cause Rubisco to preferentially bind to O2 rather than CO2. When the Rubisco enzyme binds to O2, it initiates a series of reactions that reverses the outcome of photosynthesis in a process known as photorespiration:
Photorespiration The oxidation of carbohydrates to CO2 and H2O by Rubisco, which reverses the light reactions of photosynthesis.
2 G3P → RuBP + CO2
The reverse reaction consumes energy, consumes O2, and produces CO2. This reverse reaction in plants is called photorespiration because it resembles the process of respiration. In short, what is accomplished by photosynthesis when Rubisco binds to CO2 is undone by photorespiration when Rubisco binds to O2.
The problem of photorespiration is caused in part by closed stomata. This leads to high O2 concentrations and low CO2 concentrations in the leaves. One potential solution is to keep the leaf stomata open. This would permit free gas exchange, allowing CO2 to enter the leaves and O2 to exit the leaves. This strategy works as long as plants can replace the water they also lose by transpiration when the stomata are open. However, this solution may be too costly in hot, dry environments where water is scarce. When such costs are too high, natural selection will favor traits that can either reduce the demand for water or reduce the loss of water.
C₄ Photosynthesis
We have seen that hot and dry conditions cause stomata to close, which results in a decrease of CO2, an increase of O2, and an increase in photorespiration. To address this problem, many herbaceous plants, particularly grasses growing in hot climates, have evolved a modification to the process of photosynthesis. This modification has evolved at least 45 times during the past 30 million years in at least 19 different families of angiosperms.
The C4 pathway is an adaptation that adds a step to the initial assimilation of CO2 to make it more efficient when CO2 is present at low concentrations. You can see an overview of this process in Figure 3.11. Biologists call this modification C4 photosynthesis because the first step joins CO2 with a three-carbon molecule called PEP (phosphoenol pyruvate) to produce a four-carbon molecule called OAA (oxaloacetic acid):
68
C4 photosynthesis A photosynthetic pathway in which CO2 is initially assimilated into a four-carbon compound, oxaloacetic acid (OAA).
CO2 + PEP → OAA
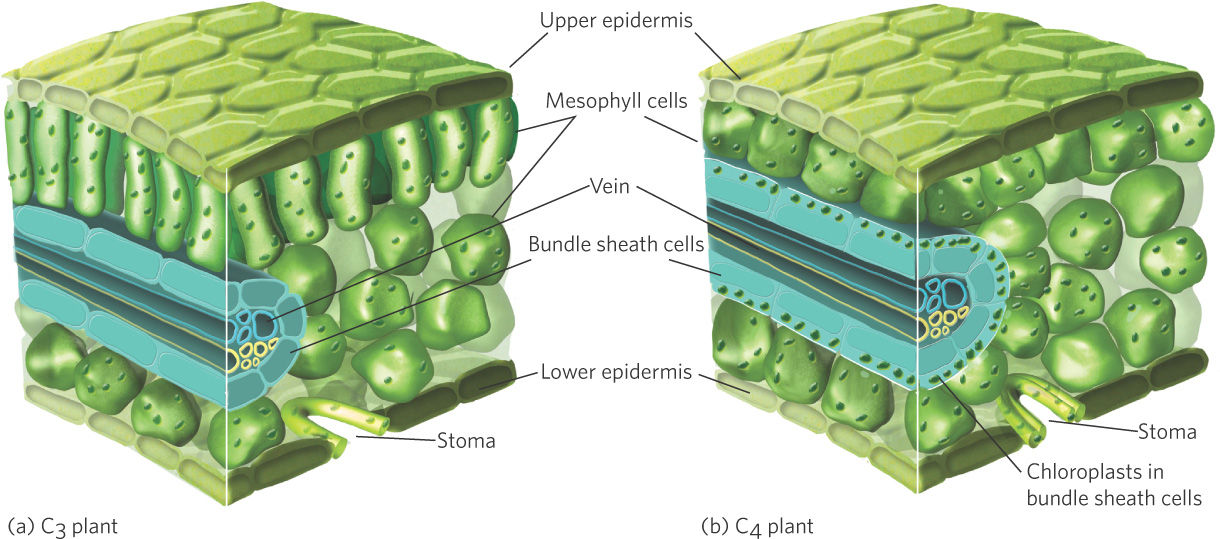
This reaction is the key difference between C3 and C4 photosynthesis. It is catalyzed by the enzyme PEP carboxylase, which has a higher affinity for CO2 than Rubisco. This additional assimilation step occurs in the mesophyll cells of the leaf, which is also the site of the light reaction. In most C4 plants, however, the Calvin cycle takes place in the bundle sheath cells that surround the leaf veins. This means that the plant must move the CO2 that has been assimilated in the mesophyll cells over to the bundle sheath cells. To do this, the plant converts OAA into malic acid, which then diffuses into the bundle sheath cells, where another enzyme breaks it down to produce CO2 and pyruvate, a three-carbon compound. In the bundle sheath cells, the chloroplasts use the CO2 that is brought over from the mesophyll cells for the Calvin cycle. To complete the cycle, pyruvate moves back into the mesophyll cells where it is converted back to PEP to be used once again.
This strategy solves the problem of photorespiration by creating concentrations of CO2 in the bundle sheath cells that are three to eight times higher than is available to C3 plants. It helps that there are multiple mesophyll cells for each bundle sheath cell, thereby providing a large number of sites for CO2 assimilation that can provide CO2 to each bundle sheath cell. At this higher CO2 concentration, the Calvin cycle operates more efficiently. In addition, because the enzyme PEP carboxylase has a high affinity for CO2, it can bind CO2 at a lower concentration in the cell. This pathway allows the stomata to remain partially or completely closed for longer periods of time, which reduces water loss. However, C4 photosynthesis has two disadvantages that reduce its efficiency: less leaf tissue is devoted to photosynthesis, and some of the energy produced by the light reactions is used in the initial C4 carbon assimilation step.
Whereas C3 plants are favored in cool, moist climates, C4 plants are favored in climates that are hot or have less abundant water. When water is abundant, the C4 pathway does not present a distinct advantage because the costs of the C3 pathway are relatively low. When water is less abundant, however, the C4 pathway has an advantage. Only about 4 percent of all plant species on Earth are C4 plants and they are primarily found in two types of nonwoody plants: grasses and sedges. C4 plants dominate tropical and subtropical grasslands and are important components of the plant communities found in arid regions of the world, including the Great Plains of North America. Plants using the C4 pathway also include many of our most important crop plants such as corn (maize), sorghum, and sugarcane. These plants are highly productive during hot growing seasons. In fact, while only 4 percent of all plant species use the C4 pathway, these plants account for 20 to 30 percent of all CO2 fixation and 30 percent of all grain production. As a result, C4 plants can play substantial roles in the ecosystems in which they live.
69
CAM Photosynthesis
Crassulacean acid metabolism (CAM) A photosynthetic pathway in which the initial assimilation of carbon into a four-carbon compound occurs at night.
Certain succulent plants that inhabit water-stressed environments, for example cacti and pineapple plants, use the same biochemical pathways as C4 plants. However, rather than separating the steps of CO2 assimilation and the Calvin cycle in space with mesophyll and bundle sheath cells, these succulent plants separate the steps in time. Plants that follow this pathway, known as crassulacean acid metabolism, or CAM,open their stomata for gas exchange during the cool night, when transpiration is minimal, and then conduct photosynthesis during the hot day. The discovery of this arrangement was first made in plants of the family Crassulaceae (the stonecrop family)—which includes the jade plant (Crassula ovata).
Like the C4 plants, CAM plants use an initial step of assimilating CO2 into OAA, which is then converted to malic acid and stored at high concentrations in vacuoles within the mesophyll cells of the leaf. The enzyme responsible for the assimilation of CO2 works best at the cool temperatures that occur at night, when the stomata are open. During the day, the stomata close, and the stored organic acids are gradually broken down to release CO2 to the Calvin cycle. A different enzyme with a higher optimum temperature, geared to promote daytime photosynthesis, regulates the regeneration of PEP from pyruvate following the release of CO2. Because CAM plants can conduct gas exchange during the night when the air is cooler and more humid, a plant using CAM photosynthesis reduces its water loss. Thus, CAM photosynthesis is an adaptation that results in extremely high water use efficiencies and enables plants that use this pathway to live in very hot and dry regions of the world. While CAM allows photosynthesis to occur in water-limited conditions, it happens at a relatively slow rate. As a result, CAM plants typically grow much more slowly than C3 or C4 plants. Figure 3.12 compares the three alternative photosynthetic pathways.
Plants possessing the C3 pathway are better adapted to cool, wet conditions, whereas plants possessing the C4 and CAM pathways are better adapted to warm and arid conditions. However, there is not a clear distinction in where these different types of plants grow. For example, regions that are hot and dry during the summer can be cool and moist during the winter and spring. As a result, such an area can be dominated by short-lived C4 plants in the summer and short-lived C3 plants in the winter and spring. Moreover, different photosynthetic pathways represent just one of several adaptations that plants have evolved to handle hot temperatures and a scarcity of water. As we will discuss in the next section, many plants also have evolved structural adaptations.
70
71
Structural Adaptations to Water Stress
Heat- and drought-adapted plants have anatomical and physiological modifications that improve water uptake and retention, reduce transpiration, and reduce the buildup of heat in their tissues. These adaptations include roots that can take advantage of different water sources, resistance to heat buildup, vein configurations that protect against air blockages, and morphological adaptations in leaves.
Plants living in arid regions often have very shallow or very deep roots, which represent two different adaptive strategies. Plants with very shallow roots, such as many species of cacti, are able to rapidly uptake water from brief rain events in which the rain does not penetrate very far into the soil. Cacti often pair this adaptation with thick, succulent tissues that can hold a great deal of water whenever it becomes available. In contrast to the cacti, some perennial shrubs, such as mesquite, have roots that can extend several meters down into the soil, allowing the shrubs to access water that is very deep below the surface.
Another strategy to combat the effects of heat and drought is to protect plant surfaces from direct sunlight with leaf resins, waxy cuticles, spines, and hairs. You can see many of these adaptations illustrated in Figure 3.13. Resins help seal off much of the leaf from water loss, whereas waxy cuticles help make the surfaces of the plant more resistant to losing water. Spines and hairs produce a boundary layer of still air that traps moisture and reduces evaporation. In some cases, to reduce water loss, the stomata are recessed into deep pits that contain hairs. Because thick boundary layers can also retard heat loss, hair-covered surfaces are also prevalent in arid environments that are cool. Long spines can also serve as structures that dissipate excess heat away from the plant.
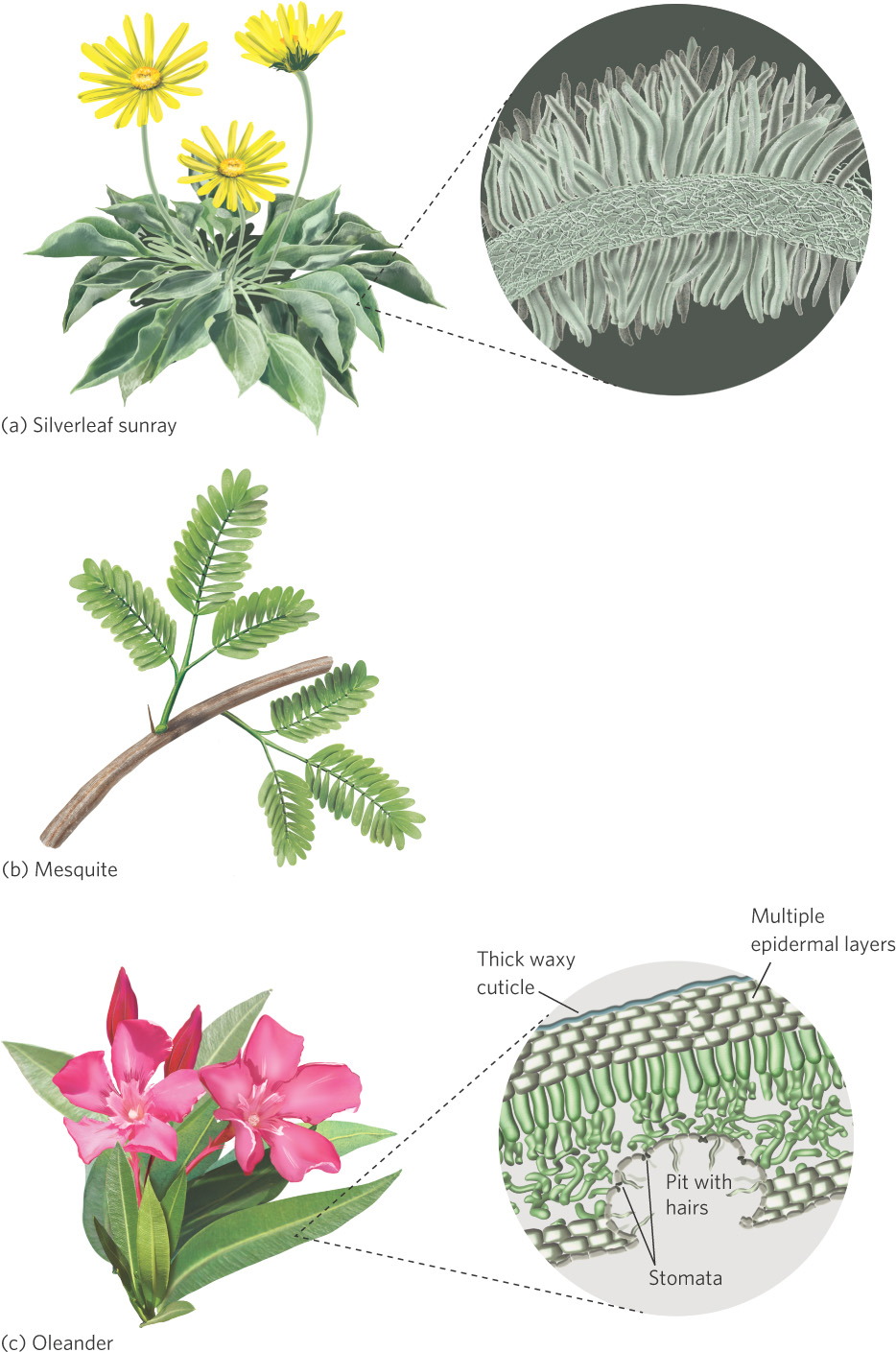
Some adaptations reduce heat buildup. Plants can reduce their heat loads by producing finely subdivided leaves with a large ratio of edge to surface area. This large amount of leaf edge breaks up the boundary layer surrounding the leaf, which helps dissipate heat from the leaf. Some desert plants have no leaves at all. Many cacti rely entirely on their stems for photosynthesis; their leaves are modified into thorns for protection.
For a long time scientists observed that plants in habitats with high temperatures generally had smaller leaves than plants in habitats with abundant water. Scientists hypothesized that the smaller leaves represented an adaptation that permitted the dissipation of heat. However, small leaves are not only found in plants from hot, dry places, but also in plants from cool, dry places. Smaller leaves also contain a higher density of large veins that transport and distribute water to the many small veins. In 2011, an international team of scientists discovered that having small leaves with a high density of large veins is actually an adaptation to overcome the problem of air bubbles, known as embolisms, which can form in large veins. Under severe drought stress, air can move into the stomata and up into the large veins, causing a bubble to form in the vein and blocking water movement. A large density of veins allows the plant to get around this problem by sending water through adjacent veins. This suggests that the small leaf size is actually an adaptation to scarce water in both hot and cool environments, and the fact that small leaves can better dissipate heat in the hot environments may be a valuable secondary benefit. Collectively, these structural adaptations help make it possible for plants to live in regions of the world that have high temperatures or scarce water.