Many organisms have evolved adaptations to variation in enemies, competitors, and mates
Many types of environmental variation can induce phenotypic plasticity. Among biotic environments, three of the best-studied types of environmental variation involve the occurrence of enemies, competitors, and mates.
Enemies
Because enemies—including predators, herbivores, parasites, and pathogens—pose a major risk to those organisms that are partially or entirely consumed, we would expect that many organisms have evolved defenses against their enemies. Like the treefrog tadpoles, many aquatic animals including fish, salamanders, insects, zooplankton, and protists alter their growth and change shape in response to predators in their environment. These changes might improve a prey’s ability to escape, make the prey difficult to fit in the predator’s mouth, or deter consumption by producing sharp spines.
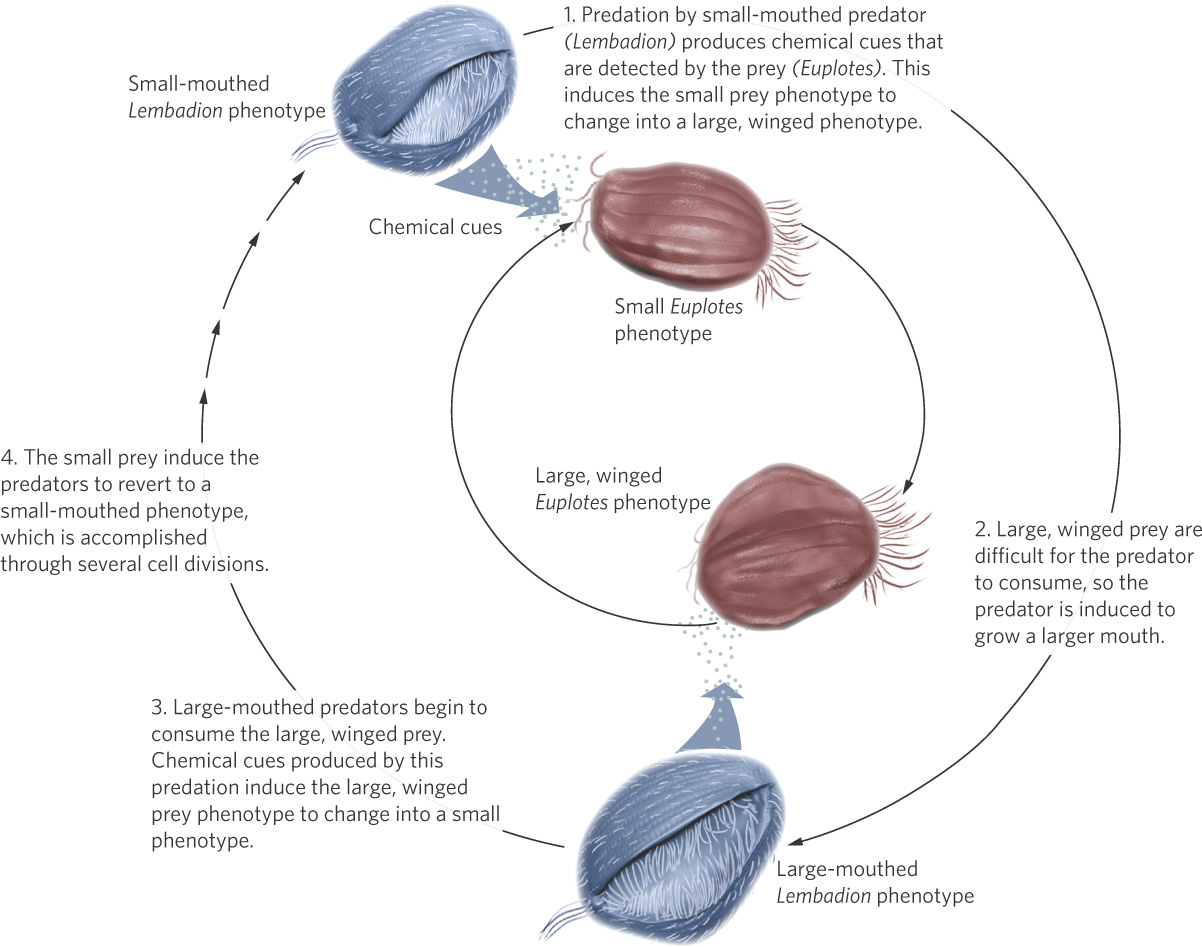
Ciliates in the genus Euplotes are tiny protists that live in lakes and streams. As you can see in Figure 4.5, these tiny organisms can sense chemical cues emitted by predators and, within hours, respond to the smell of predators by growing “wings” and numerous other projections that make them up to 60 percent larger. The larger size makes it difficult for a predator to fit Euplotes into its mouth, so the winged phenotype suffers less predation. However, because of the considerable amount of energy required to grow these projections, this phenotype takes 20 percent more time to develop. Thus, in a predator-free environment, the superior strategy for Euplotes is to have the nonwinged phenotype.
93
How do predators respond when prey make themselves more difficult to catch or consume? It turns out that the predators have plastic abilities as well. For example, when the predatory ciliate Lembadion is surrounded by the predator-induced, winged Euplotes, Lembadion grows a larger body with a bigger mouth that can engulf the winged prey. In doing so, Lembadion benefits by consuming the prey. However, once most of the large prey have been consumed and only small prey remain, the large Lembadion is poorly suited to eat the small prey. As a result, the large Lembadion experiences lower fitness than the small Lembadion. When this occurs, a large Lembadion can go through several cell divisions and revert back to having a smaller phenotype with the smaller mouth; in doing so, it is better suited to eat the small prey.
Prey also use behavioral defenses against predators. They may move away from areas containing predators or become less active to avoid detection. Some animals also congregate in refuges that are safe from predation. Although these behaviors usually reduce the risk of predation, increased safety comes at a price. When prey become less active or congregate in refuges, they spend less time feeding. Also, the supply of food in and around crowded refuges can be depleted quickly. As a result, behavioral defenses commonly come at the cost of slower growth, development, or reproduction. In the absence of predators, prey become more active and leave the refuges to find more food; this change in behavior allows more rapid growth.
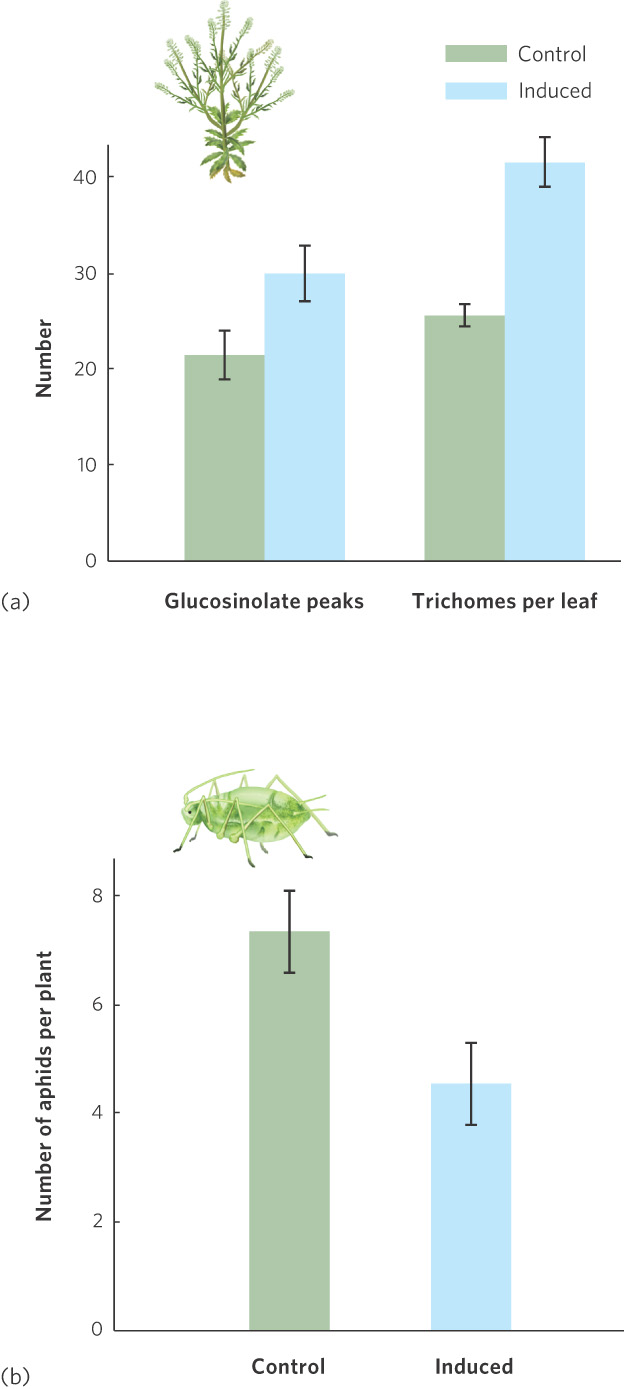
Plants also have the ability to respond to the presence of organisms that consume them. The Virginia pepperweed, a member of the mustard family, is eaten by several species of herbivores, including caterpillars and aphids. As Figure 4.6a shows, when an herbivore chews on the leaves of pepperweed, the plant rapidly develops extra leaf hairs, called trichomes, that make it harder for the leaves to be consumed. It also increases its production of glucosinolates, a group of chemicals that gives mustard its strong flavor and functions as a natural insecticide. If previously attacked plants and nonattacked plants are subsequently placed in a garden, the attacked plants, which have more trichomes and more glucosinolates, attract fewer herbivores and exhibit better survival, as shown in Figure 4.6b.
Competition for Scarce Resources
Most organisms face the challenge of scarce resources, which leads to competition. However, the intensity of competition varies across and within habitats. As a result, organisms have evolved a variety of phenotypically plastic strategies for high and low competition. As we would expect, responses to high competition often exhibit phenotypic trade-offs that favor the evolution of phenotypically plastic responses.
Jewelweed (Impatiens capensis), a flowering plant with beautiful orange flowers, is found in moist habitats throughout much of North America. In nature, it can grow in clumps that are either very sparse or very dense. The range of densities has an effect on the intensity of competition among the plants for sunlight. When jewelweed is shaded by other plants, it responds by elongating its stems, which allows the plant to become taller and to rise above the competing plants. If competition for light causes the jewelweed to grow taller, we might wonder why the plant doesn’t always grow tall. Researchers have discovered that elongated jewelweeds experience greater fitness in high-competition environments whereas short jewelweeds experience greater fitness in low-competition environments. Although the researchers were not able to identify the reasons why, it is clear that different phenotypes perform better in different environments, so a plastic phenotype is an effective way to gain high fitness when the intensity of competition varies over time and space.
94
Animals also respond to competition in a number of fascinating ways. For example, we might expect animals to spend more time searching for food when it is rare than when it is abundant. However, a second option is to find ways to extract more nutrients from the food that is available. One way to do this is to alter the size of the digestive tract, as we saw in the example of gray treefrogs, which respond to competition by increasing the length of their intestine. When the intestine is longer, food spends more time traveling through it and the organism can extract more nutrients from it.
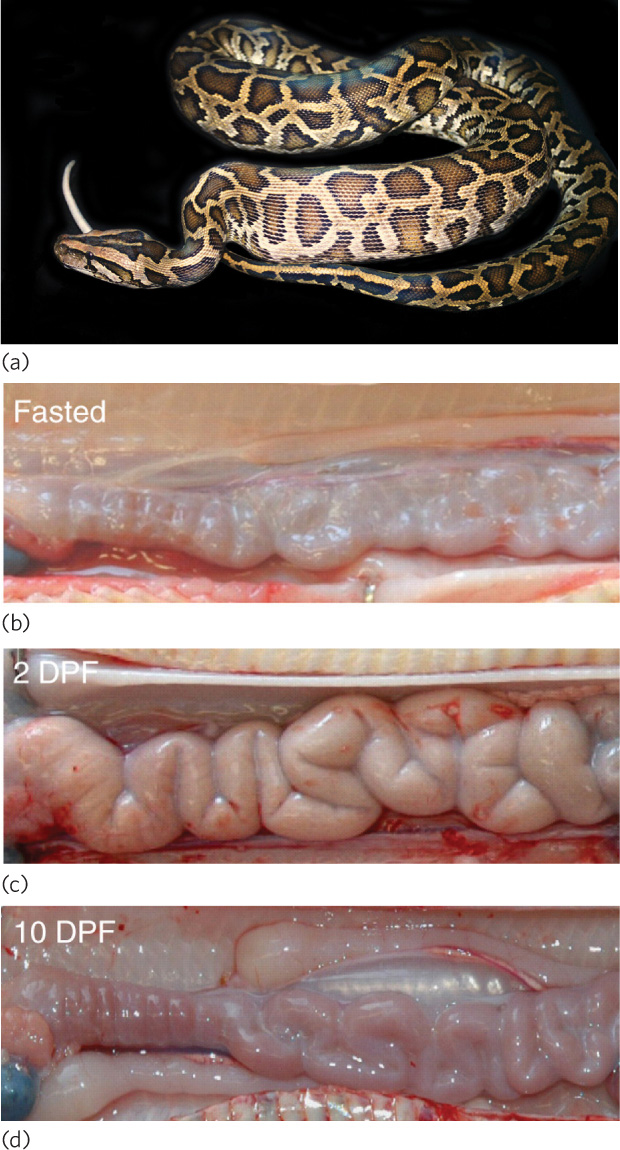
A striking example of a plastic response to variation in food availability can be found in the digestive tracts of snakes. The Burmese python, for example, can consume a large rodent that is 25 percent of its own body weight, but it may find such a prey item only once a month (Figure 4.7). Between meals, the snake’s stomach and intestine shrink a great deal, which reduces its weight and the associated carrying costs. Once a prey is consumed, however, the python can enlarge its cells and double the length of its digestive tract in just 24 hours. This dramatically increases the surface area of the intestine, which allows the snake to absorb more energy from the digested prey. The snake also sends 10 times more blood to the intestines to assist in the absorption of nutrients into the bloodstream. To handle this increase in blood flow, the snake increases the size of its heart by 40 percent, a remarkable achievement given that it occurs in 2 days.
Mates
Hermaphrodite An individual that produces both male and female gametes.
When mates are rare, reproduction can be a challenge. Consider the situation of flowering plants that depend on pollinators to deliver the pollen that contain a male’s sperm. In this situation, the probability of being pollinated can be highly variable, which means that the probability of finding a mate is also highly variable. There is a solution for flowering plants that are hermaphrodites, which are individuals that produce both male and female gametes. Such individuals are self-compatible, that is, they are capable of fertilizing their eggs with their own sperm. This process, known as self-fertilization, often carries the potential cost of inbreeding depression, in which offspring can experience reduced fitness when deleterious alleles are inherited from both the egg and the sperm. Because mate availability can be quite variable and self-fertilization can have a substantial cost, the organism must develop one of two life history choices: wait to find a mate and enjoy higher fitness, or self-fertilize. In some cases, this choice is made at the level of the entire organism. In other cases, as in many hermaphroditic species of flowering plants, the choice is made by each flower. If a flower is not pollinated within a given period of time, the flower will self-fertilize. Although self-fertilizing may result in lower fitness than breeding with a mate, low fitness is better than the zero fitness the organism would obtain by avoiding self-fertilization altogether.
Inbreeding depression The decrease in fitness caused by matings between close relatives due to offspring inheriting deleterious alleles from both the egg and the sperm.
95
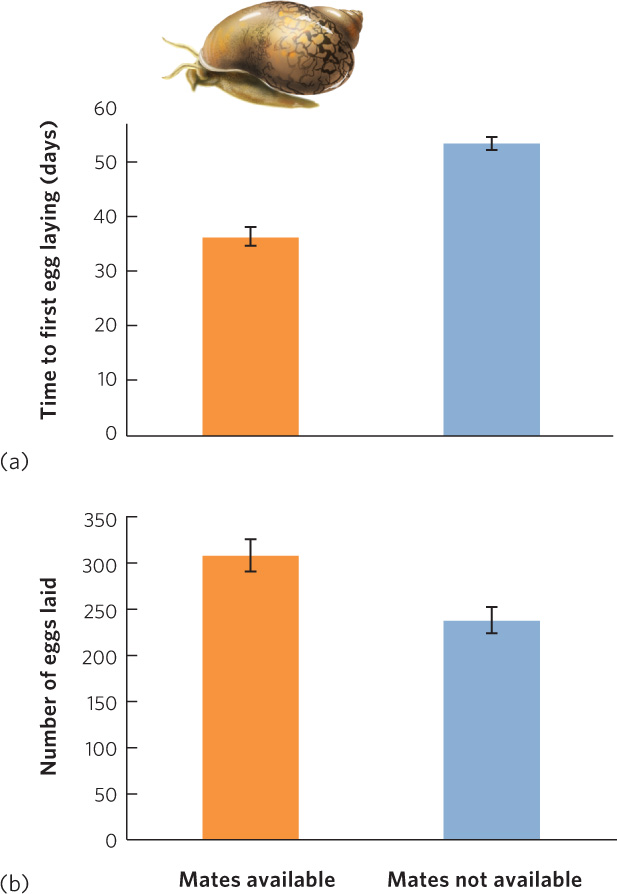
Reproductive adaptations to variation in mate availability have been studied in a variety of hermaphroditic plants and animals. For example, the common pond snail (Physa acuta) is a hermaphrodite that experiences large variation in population density, which means that it also experiences large variation in the availability of mates. When potential mates are abundant, a snail will normally mate with another individual; when potential mates are rare, it can fertilize its eggs with its own sperm. In an experiment designed to measure the fitness effects of the two alternative strategies, researchers assigned snails to one of two groups: snails living with mates or snails living without mates. The researchers then observed the time it took for each group to begin reproduction and the total number of eggs that each group of snails laid. As you can see in Figure 4.8a, snails living without mates delayed their reproduction for 2 weeks before they used the alternative strategy of self-fertilization. As predicted, this choice came at a cost. Looking at Figure 4.8b, we see that self-fertilizing snails laid many fewer eggs than snails that had mates available. However, the self-fertilizing snails did gain some fitness, which is better than deciding to forego reproduction. The good news for the pond snail, and many other hermaphroditic organisms, is that by evolving multiple strategies they can experience higher fitness over time than would be possible without a plastic strategy.