Many organisms have evolved adaptations to variable abiotic conditions
We have seen that variation in biotic conditions, including enemies, competitors, and mates, can be quite high. Abiotic conditions, including temperature, water availability, salinity, and oxygen, also vary. Faced with this abiotic variation, many species have evolved phenotypically plastic traits that allow them to improve their fitness.
Temperature
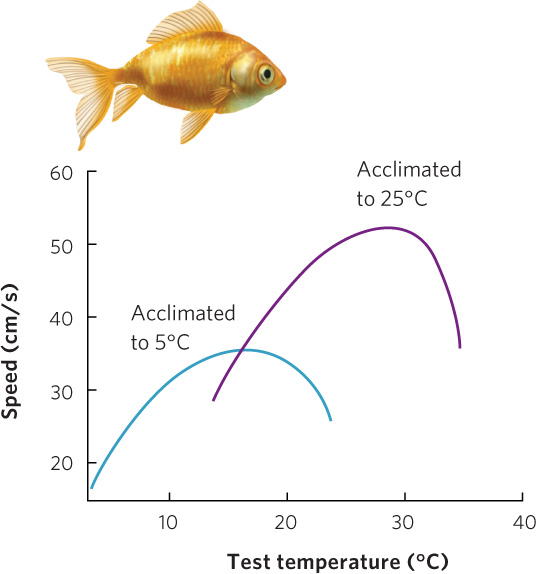
Organisms have evolved a number of plastic responses to temperature variation. As we saw in Chapter 2, isozymes in rainbow trout allow the fish to have proper nerve transmission in cold winter water and warm summer water. Isozymes are actually a form of phenotypic plasticity with a rapid response time, in some cases a matter of hours or days. For example, goldfish (Carassius auratus) can be held at either 5°C or 25°C for a few days and then tested to determine how fast they can swim in a variety of temperatures. As shown in Figure 4.9, fish acclimated to 5°C swim faster at low temperatures but swim poorly at high temperatures. In contrast, fish acclimated to 25°C swim faster at warm temperatures but swim poorly at low temperatures. This demonstrates that goldfish can adjust their physiology to maintain relatively high swimming speeds across different environmental temperatures.
Microhabitat A specific location within a habitat that typically differs in environmental conditions from other parts of the habitat.
Many animals respond to changing temperatures by moving to habitats with more favorable temperatures. Migrating birds present an extreme example; they fly every autumn to warmer latitudes. Not all animals travel a long distance. Some move to a specific location within a habitat, called a microhabitat, that contains more favorable abiotic conditions.
96
The use of microhabitats can be illustrated by the daily behavior cycle of the desert iguana (Dipsosaurus dorsalis), shown in Figure 4.10. Although the desert iguana cannot regulate its temperature by generating heat metabolically, it is able to take advantage of sunny and shady microhabitats to alter its temperature. The iguana has a preferred temperature range of 39°C to 43°C. In the southwestern United States, the air temperature can reach 45°C. As temperatures rise during the day, the lizard first moves to the shade of plants or rocks and then to a cooler underground burrow. If temperatures begin to cool, the iguana can move out of its burrow and bask in the sun to raise its temperature to its preferred range. Behavioral plasticity allows the iguana to remain within its preferred temperature range for a greater proportion of the day. In effect, the plasticity of the iguana’s behavior permits homeostasis in its body temperature.

97
Water Availability
When faced with changes in water availability, most animals can move among different microhabitats. However, plants are typically rooted in a single location and therefore face a tremendous challenge in locating water. As a result, plants have a number of phenotypically plastic adaptations for coping with water variability. Closing the stomata is one of the most common adaptations. As we saw in Chapter 3, when water is plentiful, the guard cells in a plant’s leaves open and transpiration occurs through the stomata. However, this process causes water loss. When water is scarce, these cells change shape and the stomata close to conserve water. In this way, the plant can transpire when water is abundant but stop transpiring when water is in short supply.
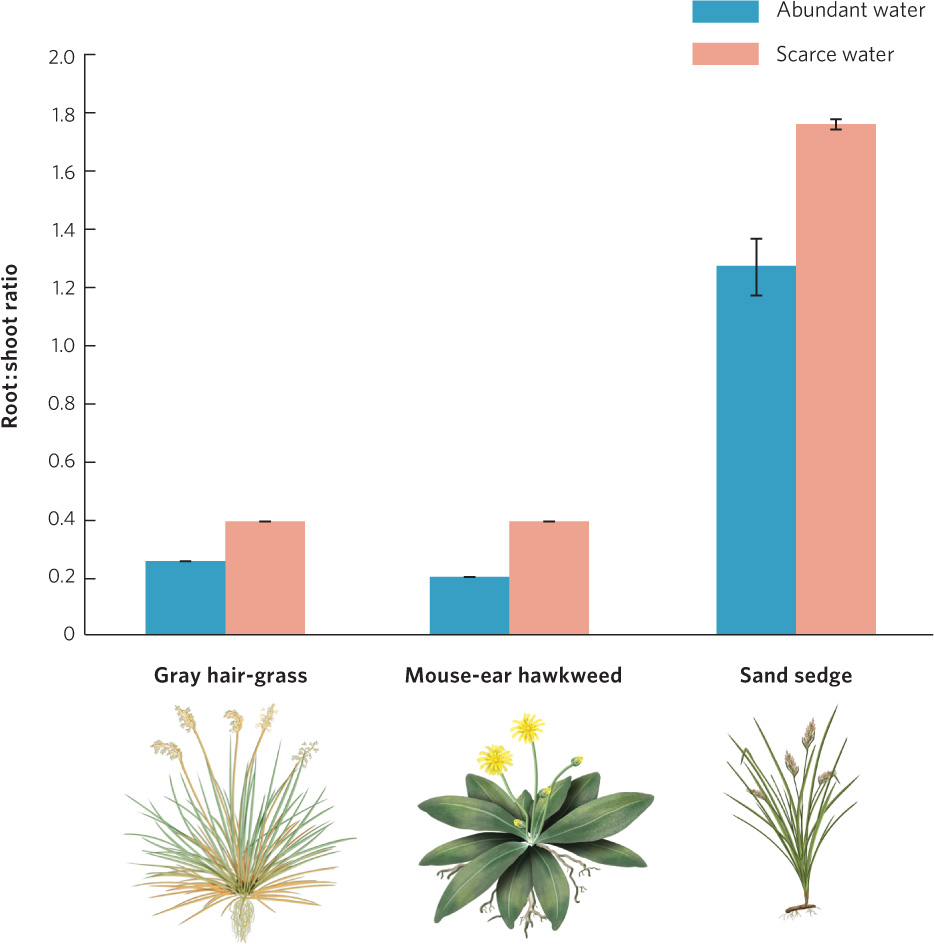
Some growth strategies of plants in response to a lack of water are even more dramatic. For example, plants living on coastal dunes in Europe commonly experience drought because water drains quickly through the sandy substrate. Three common plants of these coastal dunes—gray hair-grass (Corynephorus canescens), mouse-ear hawkweed (Hieracium pilosella), and sand sedge (Carex arenaria)—demonstrate how plants adjust their relative allocations of energy and materials to grow either roots or shoots. To demonstrate this phenomenon, researchers grew each of the three plants under conditions of abundant versus scarce water. After 5 months of growth, the researchers measured the ratio of root growth to shoot growth. Figure 4.11 shows the result. When water was abundant, the plants devoted more energy to the growth of shoots, which function primarily to photosynthesize. When water was scarce, the plants devoted more energy to the growth of roots, which expanded their ability to capture what little water was available. Given that these plants experience variation in water availability, it is clear that no single allocation strategy would be as beneficial as the phenotypically plastic strategy that they exhibit.
98
Salinity
In Chapter 2 we learned that freshwater and saltwater organisms have evolved numerous adaptations to handle their aquatic environments. However, some organisms live in aquatic environments characterized by solute concentrations that fluctuate widely over short periods of time. To survive, these organisms must have the ability to make rapid physiological adjustments. For example, the copepod Tigriopus lives along rocky Pacific coasts in pools that receive seawater infrequently from the splash of high waves (Figure 4.12). As water in the pools evaporates, the salt concentration rises to high levels, but a heavy rainfall can lower the salt concentration—a rapid reversal of environmental conditions for the copepod.

Like sharks and rays, Tigriopus manages its water balance by changing the osmotic potential of its body fluids. When the salt concentration in a pool is high, individuals synthesize large quantities of certain amino acids such as alanine and proline. These small molecules increase the osmotic potential of the body fluids to match that of the environment without the deleterious physiological consequences that come with high levels of salts or urea. However, this response to excess salts in the environment is costly in terms of the energy it requires. When individual Tigriopus are switched from 50 percent seawater to 100 percent seawater, the respiration rate of the copepods initially declines, owing to salt stress, and then increases as they synthesize alanine and proline to restore their water balance. When switched from 100 percent seawater back to 50 percent seawater, the copepods’ respiration rate immediately increases as they rapidly degrade and metabolize excess free amino acids to reduce their osmotic difference to be more in line with their new environment.
Oxygen
If you have ever been to a location at a high elevation, you probably experienced the challenge of low oxygen pressure. As you continue to rise above sea level, the air pressure becomes lower, which reduces the amount of available oxygen. At thousands of meters above sea level, breathing becomes labored and physical activity is very difficult. Animals that travel up and down mountains as part of their daily or seasonal movements, for example the llamas of South America, are able to adjust their physiology to this variation in oxygen concentration. Similarly, human mountaineers who tackle Mount Everest experience an extreme oxygen challenge for which their bodies can only partially adjust. At its peak, the mountain is 8,848 m above sea level and the oxygen pressure is only one-third of that found at sea level. To acclimate to the low oxygen conditions, climbers stop periodically for several days along the way. Initial changes include more rapid breathing and an increased heart rate. After a week or two, additional changes improve the body’s ability to carry oxygen, including an increase in the number of red blood cells and an increased concentration of hemoglobin in the red blood cells. At the highest altitudes, where humans rarely spend much time, the body is not able to fully adjust to such low oxygen concentrations; this has contributed to the deaths of many climbers at high altitudes. When climbers later return to low altitudes, their physiological changes slowly revert to their original state.
99