Organisms differ in the number of times that they reproduce, but they eventually become senescent
The number of times an individual reproduces during its lifetime varies a great deal among species. But in almost all species, individuals eventually experience a decline in body condition followed by death. For species that breed once, physiological decline and death follow rapidly. For species that breed many times, the decline in physiology comes more gradually. An interesting exception is in the case of bacteria, in which individuals do not always die. Instead, they undergo fission whereby a single-celled organism divides into two single-celled organisms. In this section we will examine the life history strategies of breeding once versus breeding several times, and investigate the causes of a gradual decline in physiology.
Semelparity and Iteroparity
Semelparity When organisms reproduce only once during their life.
Iteroparity When organisms reproduce multiple times during their life.
Annual An organism that has a life span of one year.
Organisms can evolve to reproduce only once, a phenomenon known as semelparity, or multiple times throughout their life, a phenomenon known as iteroparity. Semelparity and iteroparity do not tell us whether an organism’s life history is annual, meaning a life span of one year, or perennial, referring to a life span of more than one year. For example, organisms that live for only one year can undertake more than one episode of reproduction, or even prolonged continuous reproduction, during that time. Iteroparity is a common life history that occurs in most species of birds, mammals, reptiles, and amphibians. Semelparity is relatively rare in vertebrate animals, but it occurs in insects and many species of plants. For example, semelparity is a common life history of crop plants including wheat and corn.
Perennial An organism that has a life span of more than one year.
Bamboos, Agaves, and Yuccas
Although most semelparous organisms are short-lived, the best-known cases of plants occur in the long-lived bamboos and agaves, two distinctly different groups. Most bamboos are plants of tropical or warm temperate climates and they often form dense stands. Reproduction in bamboos does not appear to require substantial preparation or resources, as in the case of salmon. But bamboos probably have few opportunities for successful seed germination. Once a bamboo plant establishes itself in a disturbed habitat, it spreads for years by vegetative growth, continually sending up new stalks until the habitat in which it germinated is densely packed with bamboo shoots (Figure 8.8a).

In many species of bamboo, breeding is highly synchronous over large areas, such that every individual produces flowers and makes seeds in the same year. After reproduction, the future of the entire population rests with the crop of seeds. Synchronous breeding may facilitate fertilization in this wind-pollinated plant group and also may overwhelm seed predators, which cannot consume such a large crop of seeds. Some species of bamboo, such as Chinese bamboo (Phyllostachys bambusoides), have a 120-year cycle of germinating, growing, and then simultaneously flowering and producing seeds.
196
In contrast to bamboo plants, most species of agave plants live in arid climates with sparse and erratic rainfall. Ranging from the southwestern United States through Central America, agaves grow as a rosette of leaves over several years, with the duration of this growth varying among species. When the plant is ready to reproduce, it grows a gigantic flowering stalk that produces a large number of seeds (Figure 8.8b). The growth of the stalk is so rapid that it cannot be fully supported by photosynthesis in the stalk and uptake of water by the roots. Instead, the nutrients and water necessary for stalk growth are drawn from the leaves, which die soon after the seeds are produced.
Yucca plants are close relatives of the agave plants. Although most yuccas are iteroparous, a few are not. For instance, in California the yucca species Yucca whipplei contains both iteroparous and semelparous individuals. Researchers have compared the two groups—known as varieties—for total production of flower, fruits, and seeds over the lifetime of the plants. The results of this study are displayed in Figure 8.9.
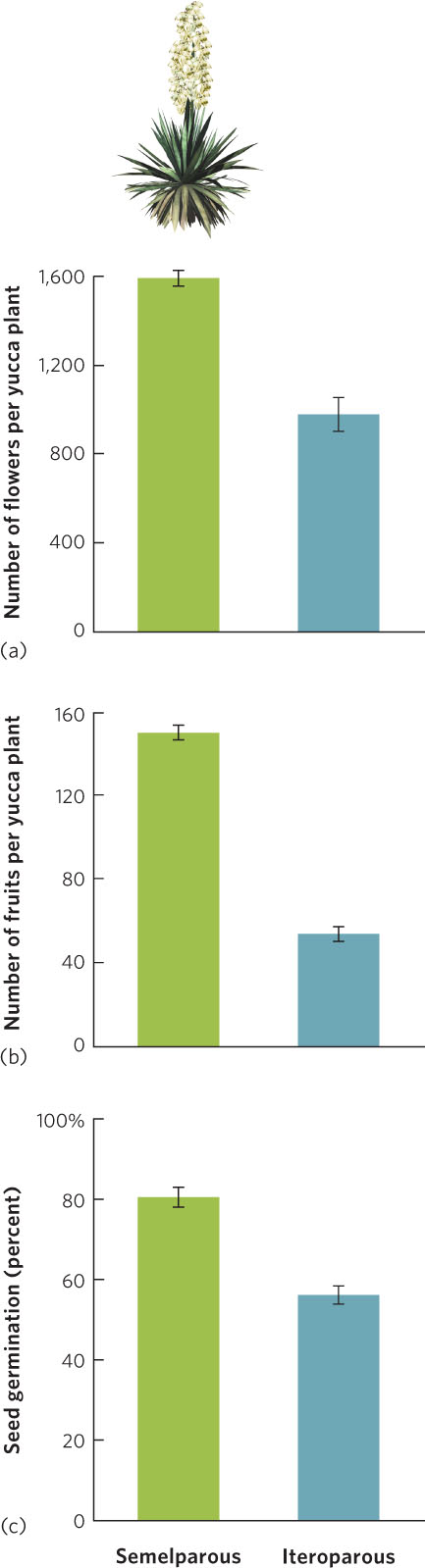
Compared to the iteroparous variety, the semelparous variety produced nearly twice as many flowers and three times as many fruits. The semelparous variety also made seeds that were more likely to germinate than those of the iteroparous variety. The iteroparous variety can reproduce earlier in life, and reproduce multiple times. The semelparous variety, which takes longer to reproduce, can produce more offspring. These differences in strategies may be related to the habitats in which the two types of individuals live. Semelparous individuals live in habitats that have higher annual precipitation and are prone to fire, which may favor a strategy of a single large reproductive effort before another fire occurs. Iteroparous individuals live along the edges of the Mojave Desert where water is much more limited and there is a reduced chance of fire. This habitat favors a strategy of multiple reproductive events over time with large amounts of reproduction during years of higher precipitation.
When we consider all of the plants and animals that practice semelparity, we see that semelparity appears to arise when there is a massive amount of energy required for reproduction, such as the long migrations of salmon and the production of giant flowering stalks of agaves and yuccas. These large energy requirements make it difficult for the individuals to survive after the reproductive event.
197
Cicadas
One of the most remarkable cases of semelparity in animals is the life cycle of periodical cicadas (Figure 8.10). Cicadas spend the first part of their life underground where they obtain nutrients from the xylem tissue of plant roots and, after some time, they emerge as adults. Their mating calls from the trees can be heard in the summer days in many parts of the Northern Hemisphere. Some cicada species have annual life cycles while other species spend several years underground with a fraction of them emerging each summer. Periodical cicadas, however, are different. They live as nymphs underground for 13 or 17 years and then emerge from the ground in synchrony to mate. The emergence of the periodical cicada is marked by nearly deafening noise as the males call to attract females during their brief mating period. The long life cycle gives the larvae time to grow to adulthood on a diet of low nutritional quality. The synchrony is probably a mechanism to overwhelm potential predators. Most of the occasional individuals that fail to emerge in synchrony come out a year earlier or later and are grabbed by predators attracted to the loud mating calls.

Scientists long wondered how periodical cicadas know when to emerge from the ground. They speculated that the cicada might count the years by the warming and cooling of the soil, or by the physiological cycles of their hosts. Researchers performed a clever experiment by growing 17-year periodical cicadas on peach trees that had been artificially selected to drop their leaves and flower twice each year. The cicadas emerged after 17 fruiting seasons had passed, rather than 17 years, demonstrating that the cicadas are sensitive to the reproductive cycles of their hosts rather than annual physical changes in their environments. How they count to 17 remains a mystery.
Senescence
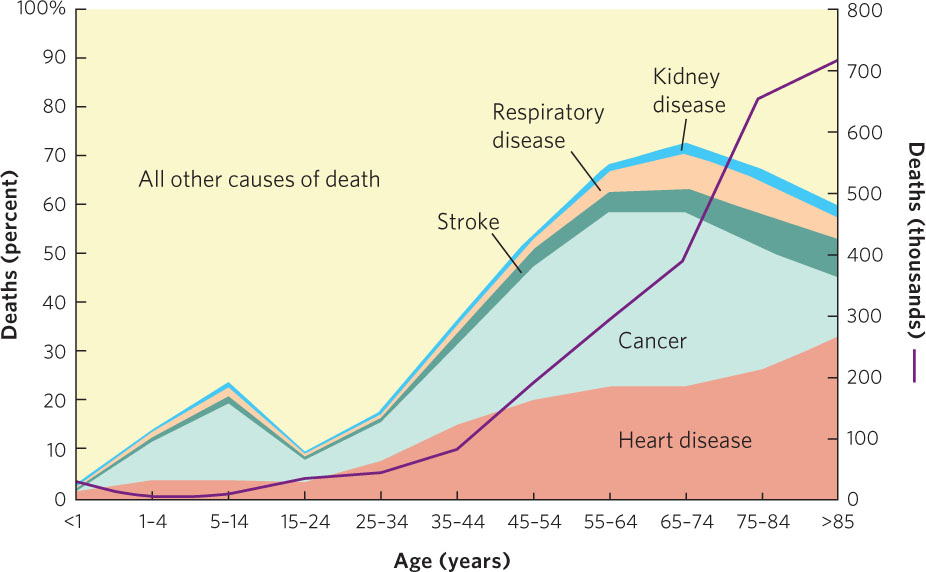
A few long-lived semelparous organisms die immediately after reproduction. However, iteroparous organisms experience a gradual deterioration in their physiological function over their lifetime. This leads to a gradual decrease in fecundity with an increase in the probability of mortality, a phenomenon known as senescence. Humans are an example of an organism that experiences senescence. Most physiological functions in humans decrease between the ages of 30 and 85 years; for example, the rate of nerve conduction and basal metabolism decrease by 15 to 20 percent, the volume of blood circulated through the kidneys decreases by 55 to 60 percent, and maximum breathing capacity decreases by 60 to 65 percent. Over time the function of the immune system and other repair mechanisms also declines. Using data from the United States population in 2007, Figure 8.11 illustrates that the incidence of death from cancers and cardiovascular diseases rises sharply with age. Birth defects in offspring and infertility also occur with increasing prevalence in women after 30 years of age, and fertility decreases dramatically in men after 60 years.
Senescence A gradual decrease in fecundity and an increase in the probability of mortality.
198
If maintaining high survival and reproduction would increase an individual’s fitness at any age, why does senescence exist? Studies of aging in a variety of animals demonstrate that senescence is an inevitable consequence of natural wear and tear. It is impossible to build a body that will not wear out eventually, just as it is impossible to build an automobile that will not wear out. Senescence might simply reflect the accumulation of molecular defects that fail to be repaired. For example, ultraviolet radiation and highly reactive forms of oxygen break chemical bonds, macromolecules become inactivated, and DNA accumulates mutations. However, this wear and tear cannot be the entire explanation for patterns of aging because maximum longevity varies widely even among species of similar size and physiology. For instance, many small insectivorous bats live 10 to 20 years in captivity, whereas mice of similar size rarely live beyond 3 to 5 years.
The rate of wear can be modified by a variety of physiological mechanisms that either prevent or repair damage. One major difference among groups that differ dramatically in life span is the cellular mechanisms for reducing the production of reactive forms of oxygen and for repairing damaged DNA and protein molecules. These appear to be better developed in long-lived animal species than in their short-lived relatives. Because these mechanisms are under genetic control, they can be modified by evolution. Mechanisms of prevention and repair require investments of time, energy, nutrients, and tissues. Therefore, the allocation of resources to these mechanisms depends on the expected life span of the individual. When a population has a low survival rate, selection should favor improvements in reproductive success at young ages and selection to delay senescence should be weak. In a population with a high survival rate, selection for delayed senescence should be strong.
This prediction is consistent with observations of natural populations. For example, because bats and birds can fly to escape predators, they lead safer lives than do rodents of a similar size. As a result, the maximum potential life spans of birds and bats are much longer than the life spans of rodents. Storm petrels, a small seabird, have a body size and metabolic rate that is similar to many rodents. However, the storm petrel can live more than 40 years whereas a rodent may only live 1 or 2 years. Because of their long life spans, birds and bats age more slowly than rodents of a similar size.