Organisms can evolve as separate sexes or as hermaphrodites
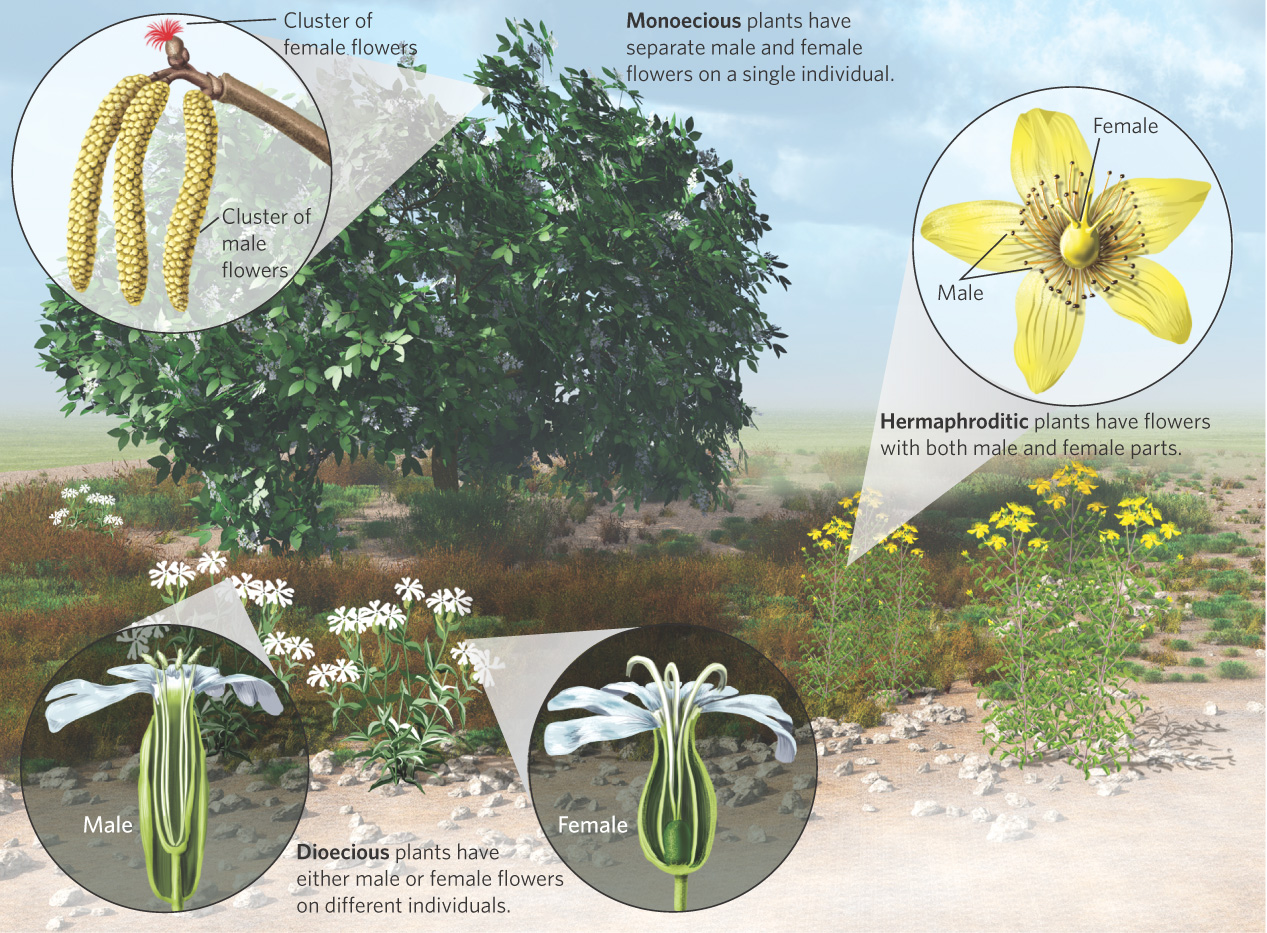
Species in nature have evolved an incredible array of sexual strategies for male and female function, as you can see in Figure 9.6. Most vertebrates and some plants have separate sexes, whereas most plants and some animals are hermaphrodites. Hermaphroditic plants, such as the wildflower known as Saint-John’s-wort (Hypericum perforatum), possess male and female functions in the same flower. Flowers that have both male and female parts within each flower are known as perfect flowers. When the two functions are produced at the same time, we call it a simultaneous hermaphrodite. Examples of simultaneous hermaphrodites include many species of mollusks, worms, and plants. When an individual possesses one sexual function and then switches to the other, we call it a sequential hermaphrodite. Some species of plants are sequential hermaphrodites, as are some mollusks, echinoderms, and fishes.
Perfect flowers Flowers that contain both male and female parts.
Simultaneous hermaphrodites Individuals that possess male and female reproductive functions at the same time.
Sequential hermaphrodites Individuals that possess male or female reproductive function and then switch to possess the other function.
Monoecious Plants that have separate male and female flowers on the same individual.
214
Dioecious Plants that contain either only male flowers or only female flowers on a single individual.
Some plants possess separate male and female flowers. Plants that have separate male and female flowers on the same individual plant are known as monoecious. For example, every hazelnut tree (Corylus americana) has separate male flowers and female flowers. When an individual plant contains only male flowers or only female flowers, the species is known as dioecious. For example, the white campion (Silene latifolia) is a wildflower comprised of some individuals that produce only male flowers and other individuals that produce only female flowers. Although perfect-flowered hermaphrodites account for more than two-thirds of flowering plant species, nearly all imaginable sexual patterns are known. Populations of some plant species can be composed of a complex mixture of hermaphrodites, males, females, and monoecious individuals. In other species, individual plants produce both perfect flowers and flowers that are only male or only female.
215
Comparing Strategies
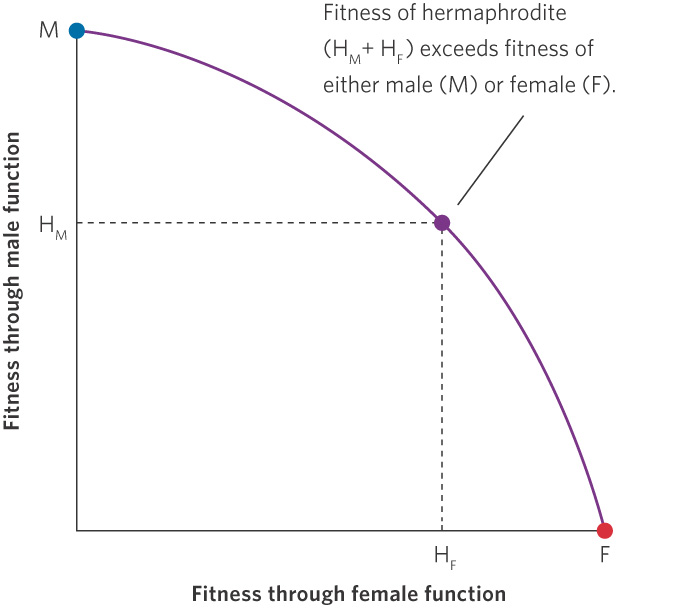
We would expect natural selection to favor the reproductive strategy with the highest fitness. For instance, in organisms such as flowering plants, a plant could evolve to make male flowers, female flowers, or hermaphrodite flowers. To determine whether evolution should favor separate sexes or hermaphrodites, we need to compare the amount of fitness an individual would gain by investing in only male or female reproduction versus the amount of fitness it would gain by investing in both male and female reproduction. As depicted in Figure 9.7, if a male individual can invest in female function and gain a lot of female fitness while only giving up a small amount of male fitness, then selection will favor the evolution of hermaphrodites. A similar scenario can be considered for a female individual that adds male function. This occurs because the total fitness as a hermaphrodite through male plus female function exceeds the fitness of being only a male or only a female. In the case of flowers, the basic flower structure and the floral display necessary to attract pollinators—for those species that rely on them—are already in place in male flowers and female flowers. This should make the fitness costs of adding a sexual function relatively small while providing large fitness benefits. As we noted earlier, about two-thirds of all flowering plant species are hermaphrodites.
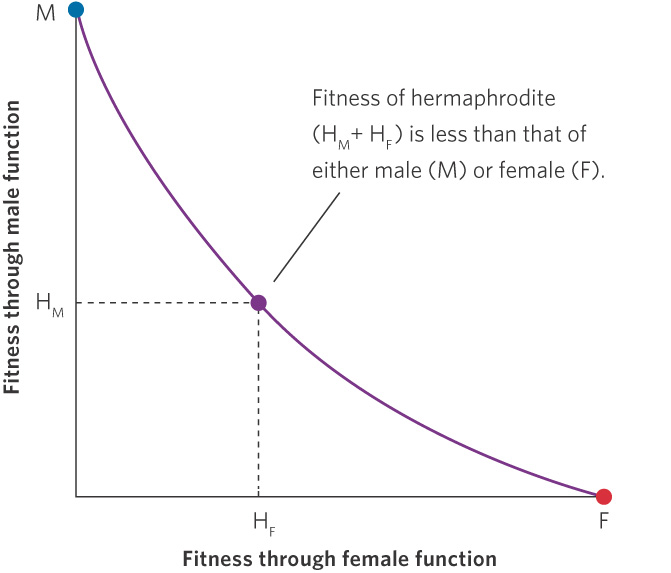
In some cases, the fitness cost of investing in a second sexual function is too high to be offset by the fitness benefits of being a hermaphrodite. Figure 9.8 illustrates how a reduction in female function can allow an investment in male function, but the total fitness is less than males and females would experience if they were to retain a single sexual function. For instance, sexual function in complex animals requires gonads, ducts, and other structures for transmitting gametes. Moreover, in many animals, being male requires large expenditures of time and energy for attracting mates and fighting with other males, while being female requires specializations for egg production or time and energy needed to care for the offspring. Because these costs can be quite high, we would predict that hermaphroditism should occur only rarely among animal species that actively seek mates and engage in brood care. In contrast, we would predict that hermaphroditism should occur commonly among sedentary aquatic animals that mate by simply shedding their gametes into the water. Researchers have found evidence that supports both of these predictions.
216
Selfing Versus Outcrossing of Hermaphrodites
One of the challenges for individuals that possess both male and female function is the problem of self-fertilization. Self-fertilization, also known as selfing, occurs when an individual uses its male gametes to fertilize its own female gametes. As we discussed in Chapter 4, self-fertilization poses a fitness cost due to inbreeding depression. Therefore, selection should favor individuals that do not use selfing when they have an opportunity to breed with other individuals, a strategy known as outcrossing. Some species avoid the problems of selfing by being sequential hermaphrodites. For example, the blue-headed wrasse (Thalassoma bifasciatum), a fish species common in coral reefs, can be functionally female when it is a small adult but then becomes functionally male later in life when it has grown larger. Similarly, if a plant releases the pollen in its anthers before making its stigma receptive to pollen, the flower will not be able to pollinate itself. Other species have self-incompatibility genes. Individuals with the same self-incompatibility genotype—including an individual mating with itself—cannot produce offspring.
Mixed Mating Strategies
As we discussed in Chapter 4, some hermaphrodites use a mixture of mating strategies. When a mate can be found, the individual prefers to breed by outcrossing to avoid the costs of inbreeding. When a willing mate cannot be found, the individual self-fertilizes. Self-fertilizing will not provide as many viable offspring as outcrossing, but it might be better than not reproducing at all.
In some cases, using a mixture of outcrossing and selfing is a response to a lack of resources. Attracting mates can be energetically expensive, as is the case with plants that must produce nectar to attract pollinators. For example, in orange jewelweed the production of outcrossing flowers is more energetically expensive than the production of selfing flowers, which do not need to invest in nectar to attract pollinators. Plants that experience herbivory of their leaves have less energy to make outcrossing flowers. As a result, these plants produce a higher proportion of selfing flowers, as shown by the data for orange jewelweed in Figure 9.9.