Sex ratios of offspring are typically balanced, but they can be modified by natural selection
In organisms with separate sexes, the sex ratio of male to female offspring is often one to one. The exceptions offer interesting insights into evolutionary forces that favor particular sex ratios in an individual’s offspring. In this section, we will examine the mechanisms that determine whether an offspring will be male or female. We will then examine the underlying reasons for the wide range of sex ratios that are observed in nature.
Mechanisms of Sex Determination
In previous chapters, we learned that the phenotypes of organisms are often determined by a combination of genetics and the environment. The sex of offspring is no different, although the influence of genetics and the environment on the sex of offspring differs among species.
Genetic Sex Determination
In mammals, birds, and many other organisms, sex is determined by inheritance of sex-specific chromosomes. In most mammals, females have two X chromosomes whereas males have one X chromosome and one Y chromosome. Birds have the opposite pattern of genetic sex determination: males have two copies of the Z chromosome whereas females have one Z chromosome and one W chromosome. In both cases, the sex that possesses two different chromosomes—male mammals and female birds—produces an approximately equal number of gametes containing each sex chromosome. On average, half the progeny in these populations will be female and half will be male.
217
In insects, genetic sex determination is accomplished in a variety of ways. In grasshoppers and crickets, for instance, all individuals are diploid, but females have two sex chromosomes whereas males have only one. In honeybees and other members of their order including other bees, ants, and wasps, sex is determined by whether an egg is fertilized. Fertilized eggs, which receive two sets of chromosomes, become females whereas unfertilized eggs become males.
Environmental Sex Determination
Environmental sex determination A process in which sex is determined largely by the environment.
In some species, sex is largely determined by the environment in a process known as environmental sex determination. In reptiles, including several species of turtles, lizards, and alligators, the sex of an individual is determined by the temperature at which the egg develops. In turtles, embryos incubated at lower temperatures typically produce males whereas those incubated at higher temperatures produce females. The reverse is generally true in alligators and lizards. This type of environmental sex determination is known as temperature-dependent sex determination. Because the genotype has the ability to produce multiple phenotypes, temperature-dependent sex determination is a type of phenotypic plasticity.
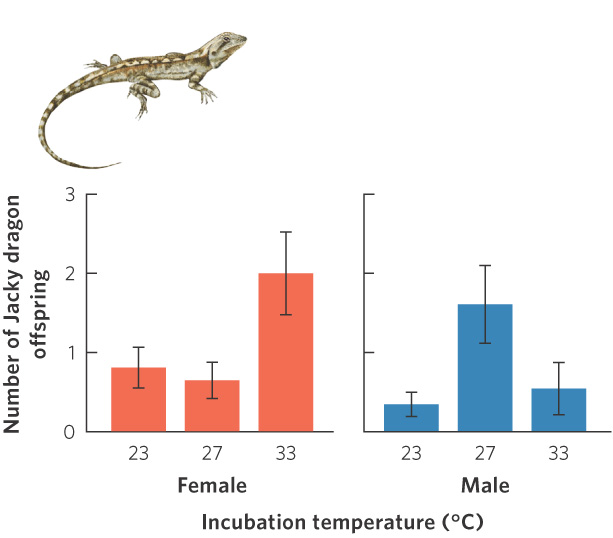
For decades, biologists have wondered whether temperature-dependent sex determination is adaptive. For example, if the temperatures that cause eggs to become male produce the most fit males, and the temperatures that cause eggs to become female produce the most fit females, temperature-dependent sex determination would be adaptive. To test this hypothesis, one would need to produce male and female offspring from eggs incubated across a range of different temperatures. However, since temperature is the very factor that determines their sex, we cannot naturally produce males and females at each temperature and compare their performance. This problem was solved in 2008 in a study of the Jacky dragon (Amphibolurus muricatus). In this lizard species from Australia, females are produced when incubated at low and high temperatures whereas both males and females are produced when incubated at intermediate temperatures. Because of this pattern, researchers could easily produce females at all three temperatures. To produce males at the highest and lowest temperatures, the researchers injected a hormone inhibitor that prevented the embryos from becoming female, thereby overriding the normal response to temperature effects. This manipulation allowed the researchers to produce male and female lizards at all three temperatures. Once the lizards hatched, the animals were put outside in field enclosures for 3 years. At the end of 3 years, the researchers determined the number of offspring produced by adults that had been incubated as eggs at different temperatures. As shown in Figure 9.10, males incubated at the intermediate temperature subsequently fathered more offspring than males incubated at the low and high temperatures. Females that incubated at the high temperature subsequently laid more eggs than females incubated at intermediate temperatures, although females incubated at low temperatures laid a similar number of eggs as females incubated at intermediate temperatures. This was one of the first studies in reptiles to demonstrate that temperature-dependent sex determination appeared to be adaptive.
Not all cases of environmental sex determination are driven by temperature. In some species, sex determination is driven by the social environment in which an individual lives. As mentioned in our discussion of sequential hermaphrodites, the blue-headed wrasse is a coral reef fish that is able to change from female to male as it gets older (Figure 9.11). Blue-headed wrasses typically begin life as females and live in large schools along with one or a few dominant males. If for some reason the dominant male leaves or dies, the largest female will then change sex and become the new dominant male. Being large is important for males, because they must defend a territory from other males that attempt to breed with the females in the school.

218
Offspring Sex Ratio
Now that we understand the mechanisms that help determine the sex of offspring, we can consider the factors that favor particular sex ratios of offspring. Biologists increasingly appreciate that a female can have a large influence on the sex ratios of her offspring. In species where the males have two different types of sex chromosomes, as in mammals, researchers have found that the females of some species can control whether X-chromosome or Y-chromosome sperm are allowed to fertilize their eggs. In species where the females contain two different types of sex chromosomes, as in birds, the females of some species can determine the sex ratio of their offspring by controlling the fraction of her eggs that receive the Z versus the W chromosome during meiosis. In the hymenopteran insects—bees, wasps, and ants—the female determines the sex of her offspring by whether or not she fertilizes the eggs.
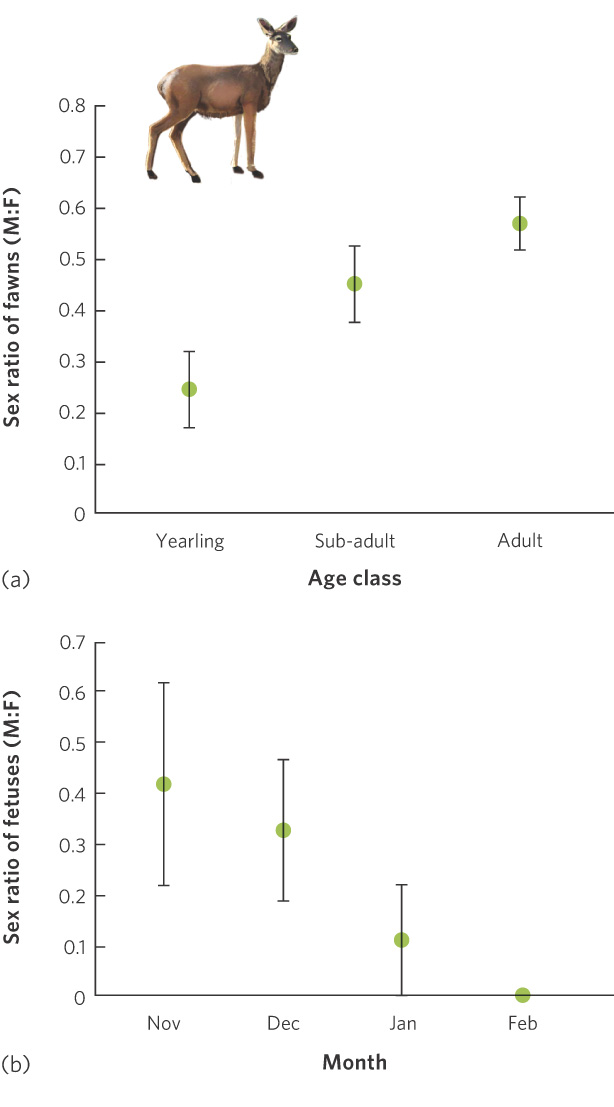
A different approach to controlling offspring sex ratio is through selective abortion. In red deer (Cervus elaphus), for example, adult females breed in early autumn and give birth the following spring. Sons are often larger at birth and require more of the mother’s milk than daughters. As a result, sons require a larger investment from the mother than daughters. Researchers in Spain examined the fetuses of 221 harvested red deer to determine whether the sex ratio of the fetuses was affected by the mother’s age—categorized as adult, subadult, or yearling—and whether the sex ratio changed through the gestation period. As shown in Figure 9.12a, adult mothers produced a nearly even offspring sex ratio. In contrast, yearling mothers, which are smaller and have less energy, were much more likely to produce the less energetically expensive daughters. Averaged across all months of the deer harvest, yearling mothers carried about 25 percent male fetuses and 75 percent female fetuses. To determine how the yearling mothers accomplish this uneven sex ratio, researchers examined yearling mothers that were harvested at different months throughout the winter, illustrated in Figure 9.12b. They found that the yearling mothers initially had a nearly even sex ratio of offspring. As winter progressed, however, the proportion of male fetuses declined sharply, suggesting that the yearling mothers were capable of selectively aborting the more expensive male fetuses as gestation progressed. This phenomenon is not restricted to red deer; it has also been observed in other mammals including rats and mice.
219
Regardless of how the sex ratio is controlled, in most species the sex ratio of male to female offspring is nearly even. Is this a ratio produced by default, or are there adaptive reasons for it? To answer this question we can compare the conditions that favor a one-to-one sex ratio with the conditions that favor a deviation from that ratio.
Frequency-Dependent Selection
Frequency-dependent selection When the rarer phenotype in a population is favored by natural selection.
To understand how selection results in a one-to-one sex ratio in most species, we have to consider the best strategy for an individual whenever the sex ratio might deviate from being balanced. Let’s imagine, for instance, that a population had more females than males and each male bred with only one female. In this situation, some females would remain unbred. A parent that produced all sons would have higher fitness than a parent that produced an even number of sons and daughters, because some of the daughters might not find mates. Similarly, if a population had a surplus of sons and one male bred with only one female, a parent that produced all daughters would have higher fitness than a parent that produced an even number of sons and daughters, because some of the sons might not find mates. As you can see in this example, individuals of the less abundant sex enjoy greater reproductive success because they compete with fewer individuals of the same sex for breeding. Thus, whenever the population has an abundance of one sex, natural selection will favor any parents that produce offspring of the less abundant sex. Over time, as the less abundant sex becomes more common and the common sex becomes less abundant, populations tend to be balanced at an approximate one-to-one ratio. Therefore, the best sex ratio strategy for parents depends on the frequencies of males and females in a population. The evolution of the sex ratio is said to be the product of frequency-dependent selection, which occurs when natural selection favors the rarer phenotype in a population.
Highly Skewed Sex Ratios
We have seen that natural selection favors a one-to-one sex ratio. In some cases, however, we can observe highly skewed sex ratios. Fig wasps (Pegoscapus assuetus) offer a good example of this phenomenon. The female fig wasp arrives at a flower of a fig tree carrying pollen from another fig tree. She crawls in through a small hole in the flower and pollinates the flower. Once inside, the female lays its eggs in the developing fruit and then dies. It is fascinating that her eggs can be comprised of up to 90 percent daughters. Similar to the bees that we discussed at the beginning of this chapter, wasps can easily adjust the sex ratio of their offspring because fertilized eggs become daughters whereas unfertilized eggs become sons. Once the eggs hatch, the larvae feed on the fig fruit and seeds. The larvae metamorphose into adult wasps and breed with their siblings while still inside the flower. The young males breed with the young females, chew a hole in the side of the flower, and then die. The fertilized young females then escape through the hole and fly off to pollinate a new fig flower.
ANALYZING ECOLOGY
Frequency-Dependent Selection
We can better understand frequency-dependent selection if we work though an example with real numbers. The American black vulture (Coragyps atratus) is a large bird that feeds on animal carcasses throughout much of North and South America. The female vulture typically lays two eggs in its nest.
If a population is composed of five females and two males, and we assume that a male can breed with more than one female, how many copies of genes, on average, does each male and female contribute to the next generation?
Total number of eggs = 10
Average female fitness = 10 female gene copies ÷ 5 females = 2 gene copies/female
Average male fitness = 10 male gene copies ÷ 2 males = 5 gene copies/male
If a population is composed of five females and eight males, how many copies of genes, on average, does each male and female contribute to the next generation?
Total number of eggs = 10
Average female fitness = 10 female gene copies ÷ 5 females = 2 gene copies/female
Average male fitness = 10 male gene copies ÷ 8 males = 1.25 gene copies/male
In this example, you can see that when males are the less abundant sex, males have the higher fitness. When females are the less abundant sex, females have the higher fitness.

- 1. Four males and five females
- 2. Six males and five females
Given the four scenarios that we have explored, what sex ratio will be favored by natural selection over the long term?
If there are four males and five females:
Average male fitness = 10 gene copies ÷ 4 males = 2.5 gene copies/male
Average female fitness = 10 gene copies ÷ 5 females = 2.0 gene copies/female
If there are six males and five females:
Average male fitness = 10 gene copies ÷ 6 males = 1.7 gene copies/male
Average female fitness = 10 gene copies ÷ 5 females = 2.0 gene copies/female
Over the long term, if either sex becomes rare, it will have higher fitness, which will subsequently cause it to become more common. Over the long term, this process will favor an equal sex ratio.
Local mate competition When competition for mates occurs in a very limited area and only a few males are required to fertilize all of the females.
220
The high proportion of daughters among fig wasps occurs because male competition for females is among brothers. This phenomenon, known as local mate competition, occurs when competition for mates takes place in a limited area and only a few males are required to fertilize all the females. When only one fertilized mother enters a fig flower, the only males available to fertilize her daughters are her sons. From the mother’s perspective, it does not matter which son passes along her genes to her grandoffspring. One son can fertilize multiple daughters, which means there is no fitness benefit to having multiple sons. The mother’s fitness is therefore a function of how many eggs can be produced by her daughters, so it is in the mother’s best interest to produce many daughters and just enough sons to fertilize those daughters. As you can see in Figure 9.13, when mating options are restricted such that the only mates available for daughters are their brothers, mothers that produce a higher proportion of daughters than sons will have more grandoffspring and therefore greater evolutionary fitness.
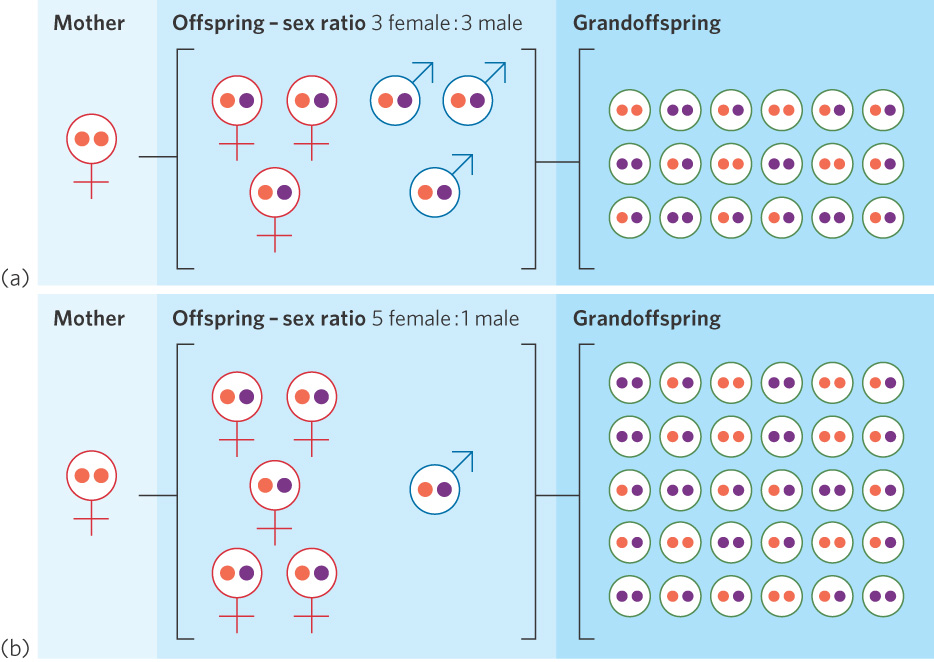
Sometimes, however, two or more females lay their eggs in the same flower. In this situation, a mother’s male offspring can mate either with their sisters or with the daughters of the other mothers. When this occurs, a mother will obtain greater fitness if she produces extra sons so she has enough sons to fertilize all the females in the flower. As expected, researchers observe that when a fig flower contains more than one mother, the mothers lay a higher proportion of male eggs.