Brain Development
neurons  cells that are specialized for sending and receiving messages between the brain and all parts of the body, as well as within the brain itself
cells that are specialized for sending and receiving messages between the brain and all parts of the body, as well as within the brain itself
cell body a component of the neuron that contains the basic biological material that keeps the neuron functioning
As you will see, the collaboration between nature and nurture takes center stage in the development of the brain and nervous system. Before discussing developmental processes in the formation of the brain, however, we need to consider the basic components of this “most complex structure in the known universe” (R. F. Thompson, 2000, p. 1).
Fundamental to all aspects of behavioral development is the development of the central nervous system and especially the brain. The brain is the font of all thought, memory, emotion, imagination, personality—in short, the behavior, capacities, and characteristics that make us who we are.
Structures of the Brain
dendrites neural fibers that receive input from other cells and conduct it toward the cell body in the form of electrical impulses
In our examination of the structures of the brain, we focus our discussion on two that are central to behavior—the neuron and the cortex, as well as some of their substructures.
Neurons
axons neural fibers that conduct electrical signals away from the cell body to connections with other neurons
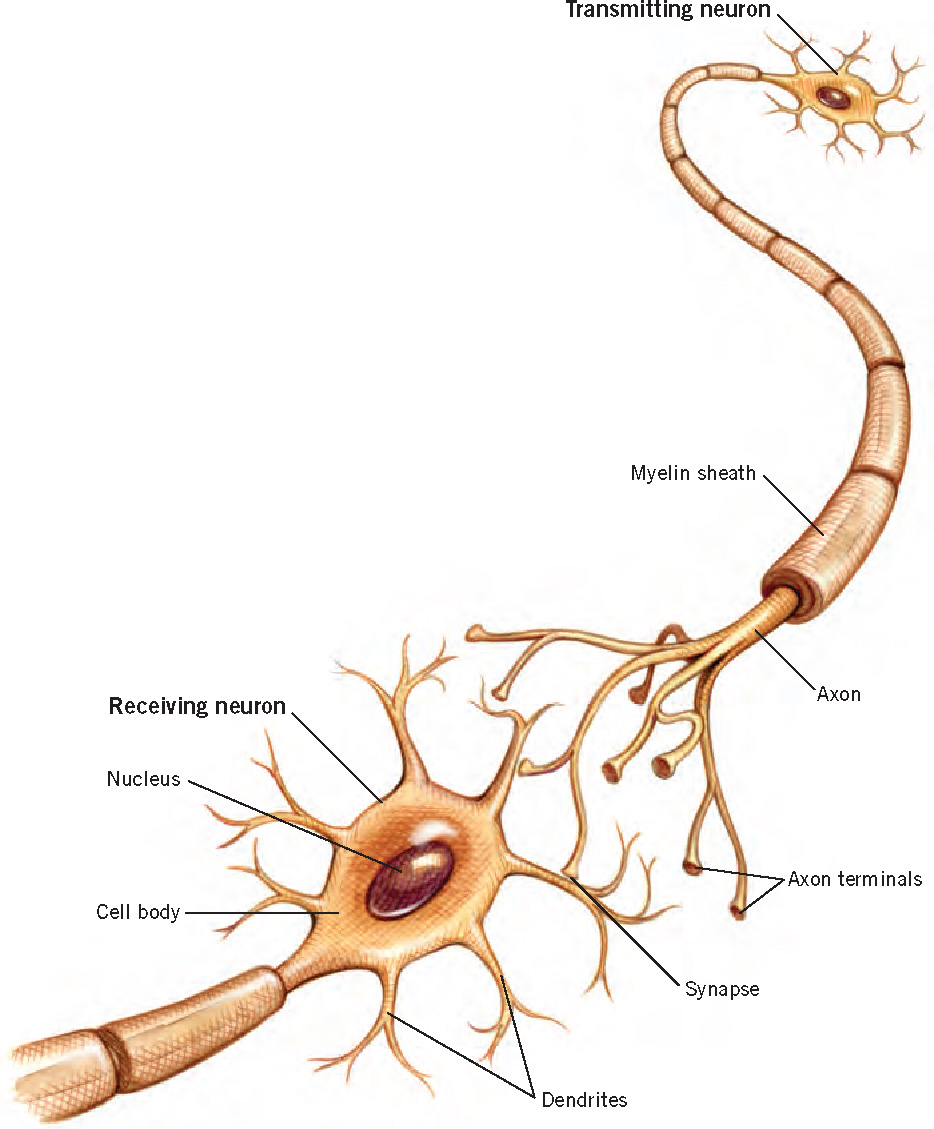
The business of the brain is information. The basic units of the brain’s remarkably powerful informational system are its more than 100 billion neurons (Figure 3.6), which constitute the gray matter of the brain. These cells are specialized for sending and receiving messages between the brain and all parts of the body, as well as within the brain itself. Sensory neurons transmit information from sensory receptors that detect stimuli in the external environment or within the body itself; motor neurons transmit information from the brain to muscles and glands; and interneurons act as intermediaries between sensory and motor neurons.
synapses microscopic junctions between the axon terminal of one neuron and the dendritic branches or cell body of another
Although they vary substantially in size, shape, and function, all neurons are made up of three main components: (1) a cell body, which contains the basic biological material that keeps the neuron functioning; (2) dendrites, fibers that receive input from other cells and conduct it toward the cell body in the form of electrical impulses; and (3) an axon, a fiber (anywhere from a few micrometers to more than a meter in length) that conducts electrical signals away from the cell body to connections with other neurons.
107
glial cells cells in the brain that provide a variety of critical supportive functions
Neurons communicate with one another at synapses, which are microscopic junctions between the axon terminal of one neuron and the dendritic branches of another. In this communication process, electrical and chemical messages cross the synapses and cause the receiving neurons either to fire, sending a signal on to other neurons, or to be inhibited from firing. The total number of synapses is staggering—hundreds of trillions—with some neurons having as many as 15,000 synaptic connections with other neurons.
Glial Cells
myelin sheath a fatty sheath that forms around certain axons in the body and increases the speed and efficiency of information transmission
Glial cells, the brain’s white matter, make up nearly half the human brain, outnumbering neurons 10 to 1. They perform a variety of critical functions, including the formation of a myelin sheath around axons, which insulates them and increases the speed and efficiency of information transmission. The importance of myelin is highlighted by the severe consequences that can arise from disorders that affect it. For example, multiple sclerosis is a disease in which the immune system attacks myelin, interfering with neuronal signaling and producing varying degrees of physical and cognitive impairment. Several psychiatric disorders, including schizophrenia and bipolar disorder, are also linked to defects in the gene that regulates production of myelin (e.g., Hakak et al., 2001; Tkachev et al., 2003).
108
cerebral cortex the “gray matter” of the brain that plays a primary role in what is thought to be particularly humanlike functioning, from seeing and hearing to writing to feeling emotion
Glial cells play a further role in communication within the brain by influencing the formation and strengthening of synapses and by communicating biochemically among themselves in a network separate from the neural network, and allowing them to efficiently regulate many aspects of brain activity (Fields, 2004).
The Cortex
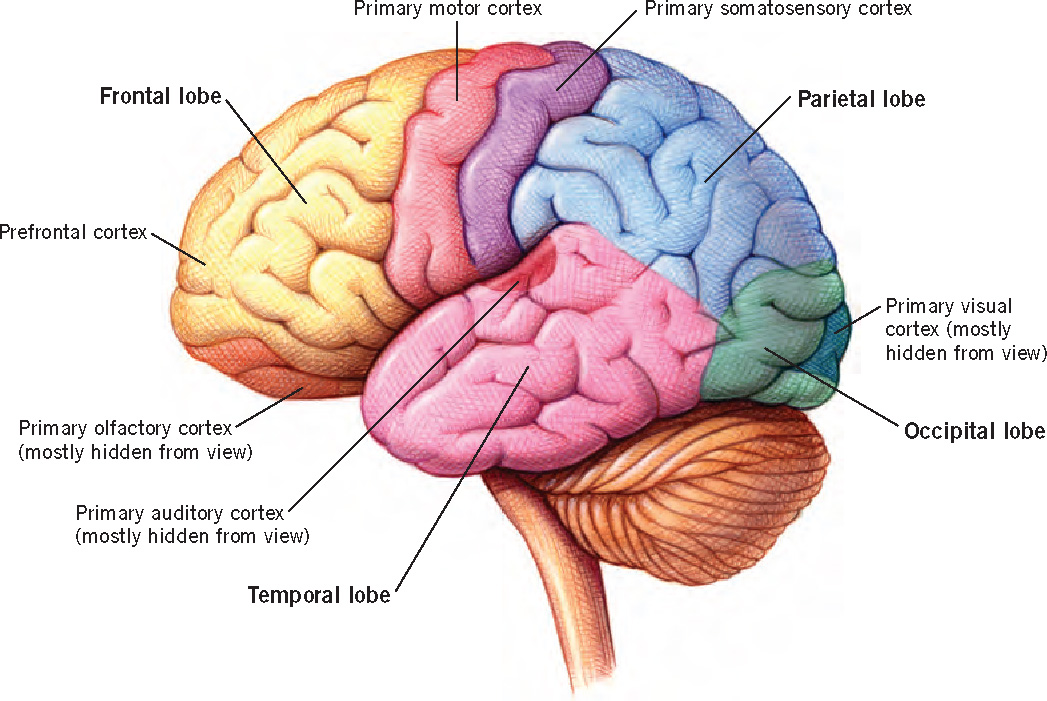
The cerebral cortex, the surface of which is shown in Figure 3.7, is considered the “most human part of the human brain” (McEwen & Schmeck, 1994). Over the course of human evolution, the brain expanded greatly in size. Almost all of this increase occurred in the cerebral cortex, which constitutes 80% of the brain, a much greater proportion than in other species. The folds and fissures that are apparent in Figure 3.7 form during development as the brain grows within the confined space of the skull; these convolutions make it possible to pack more cortex into the limited space.
lobes major areas of the cortex associated with general categories of behavior
occipital lobe the lobe of the cortex that is primarily involved in processing visual information
temporal lobe the lobe of the cortex that is associated with memory, visual recognition, and the processing of emotion and auditory information
parietal lobe governs spatial processing as well as integrating sensory input with information stored in memory
frontal lobe associated with organizing behavior; the one that is thought responsible for the human ability to plan ahead
The cortex plays a primary role in a wide variety of mental functions, from seeing and hearing to reading, writing, and doing arithmetic and to feeling compassion and communicating with others. As Figure 3.7 shows, the major areas of the cortex—the lobes—can be characterized in terms of the general behavioral categories with which they are associated. The occipital lobe is primarily involved in processing visual information. The temporal lobe is associated with memory, visual recognition, speech and language, and the processing of emotion and auditory information. The parietal lobe is important for spatial processing. It is also involved in the integration of information from different sensory modalities, and it plays a role in integrating sensory input with information stored in memory and with information about internal states. The frontal lobe, the brain’s “executive,” is involved in cognitive control, including working memory, planning, decision making, and inhibitory control. Information from multiple sensory systems is processed and integrated in the association areas that lie in between the major sensory and motor areas.
109
association areas parts of the brain that lie between the major sensory and motor areas and that process and integrate input from those areas
Although it is convenient to think of different cortical areas as if they were functionally specific, they are not. It has become increasingly clear that complex mental functions are mediated by multiple areas of the brain, with an extraordinary degree of interactivity both within and across brain regions. A given area may be critical for some ability, but that does not mean that control of that ability is located in that one area. (Box 3.3 examines some of the techniques that researchers use to learn about brain functioning.)
cerebral hemispheres the two halves of the cortex; for the most part, sensory input from one side of the body goes to the opposite hemisphere of the brain
Cerebral lateralization The cortex is divided into two separate halves, or cerebral hemispheres. For the most part, sensory input from one side of the body goes to the opposite side of the brain, and the motor areas of the cortex control movements of the opposite side of the body. Thus, if you pick up a hot pot with your right hand, it is the left side of the brain that receives the sensory response, registers the pain, and initiates the motor response to let go immediately.
corpus callosum a dense tract of nerve fibers that enable the two hemispheres of the brain to communicate
cerebral lateralization the specialization of the hemispheres of the brain for different modes of processing
The left and right hemispheres of the brain communicate with each other primarily by way of the corpus callosum, a dense tract of nerve fibers that connect them. The two hemispheres are specialized for different modes of processing, a phenomenon referred to as cerebral lateralization. There are notable similarities across species in hemispheric specialization. For example, most aspects of speech and language are lateralized to the left hemisphere in humans, with a similar asymmetry observed for communicative signals in nonhuman species from mice to primates (Corballis, 2009).
Developmental Processes
How does the incredibly complex structure of the human brain come into being? You will not be surprised to hear that, once again, a partnership of nature and nurture is involved. Some aspects of the construction of the brain are set in motion and tightly controlled by the genes, relatively independent of experience. But, as you will see, other aspects are profoundly influenced by experience.
Neurogenesis and Neuron Development
neurogenesis the proliferation of neurons through cell division
In the 3rd or 4th week of prenatal life, cells in the newly formed neural tube begin dividing at an astonishing rate—at peak production, 250,000 new cells are born every minute. Neurogenesis, the proliferation of neurons through cell division, is virtually complete by around 18 weeks after conception (Rakic, 1995; Stiles, 2008). Thus, most of the roughly 100 billion neurons you currently possess have been with you since before you were born. Notably, however, we do continue to generate new neurons throughout life. During bouts of learning, for example, neurogenesis occurs in the hippocampus, a brain region important for memory processes (E. Gould et al., 1999). Neurogenesis does not always occur, however: it can be inhibited by stress (Mirescu & Gould, 2006). This pattern of results suggests that neurogenesis later in life is not fixed and predetermined but is instead adaptive, increasing under rewarding conditions and decreasing in threatening environments (e.g., Glasper, Schoenfeld, & Gould, 2012).
110
Box 3.3: a closer look
MAPPING THE MIND
Developmental scientists employ a variety of techniques to determine what areas of the brain are associated with particular behaviors, thoughts, and feelings and how brain functions change with age. The existence of increasingly powerful techniques for investigating brain function has sparked a revolution in the understanding of the brain and its development. Here, we provide examples of some of the techniques most often used to map the mind and its workings in children.
Electrophysiological Recording
Video: Event-Related Potential (ERP) Research

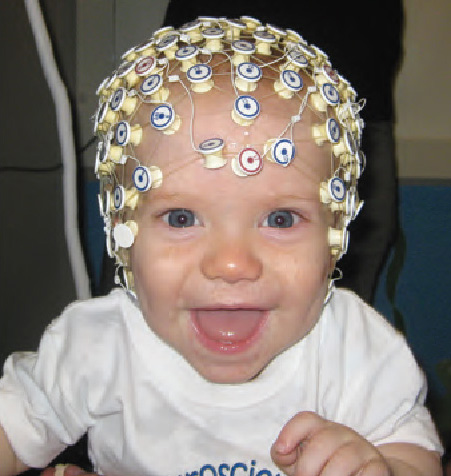
One of the techniques most often used by developmental researchers to study brain function is based on electroencephalographic (EEG) recordings of electrical activity generated by neurons. EEG is completely noninvasive (the recordings are obtained through an electrode cap that simply rests on the scalp, and can contain hundreds of electrodes), so this method can be used successfully even with infants (see photo at right). EEG recordings provide detailed information about the time course of neural events, and have provided valuable information about a variety of brain-behavior relations.
event-related potentials (ERPs) changes in the brain’s electrical activity that occur in response to the presentation of a particular stimulus
An electrophysiological technique that is very useful for studying the relation between brain activity and specific kinds of stimulation is the recording of event-related potentials (ERPs), that is, changes in the brain’s electrical activity that occur in response to the presentation of a particular stimulus. One contribution these measures have made is that they reveal continuity over time. For instance, studies of infants’ ERPs in response to native-speech sounds have shown that infants’ ability to discriminate among these sounds predicts their language growth over the next few years (Kuhl et al., 2008). A related method, called magnetoencephalography (MEG), detects magnetic fields generated by electrical currents in the brain. MEG has the added benefit of being the only noninvasive imaging method that can be used to study the fetal brain. Although the use of MEG for this purpose is in its early stages, researchers have so far been able to detect fetal neural responses to auditory stimuli and flashes of light displayed on the mother’s abdomen, as well as habituation responses to repeated stimuli (Sheridan et al., 2010).
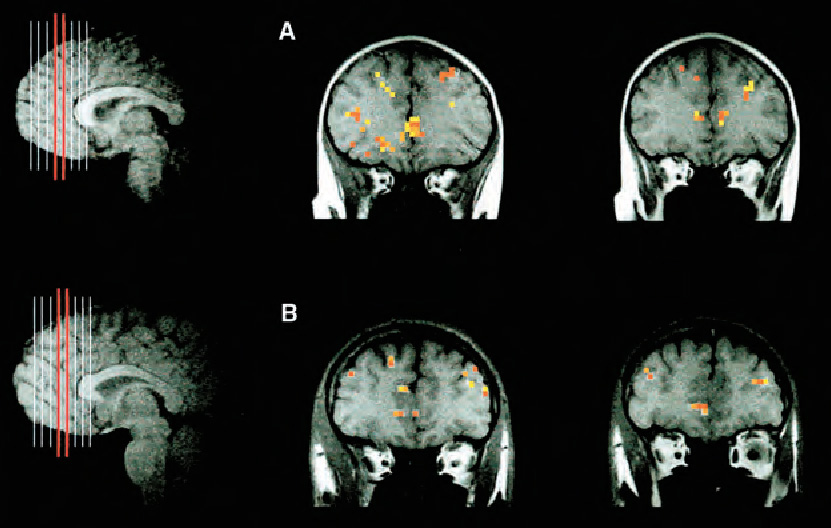
Functional Magnetic Resonance Imaging
Functional magnetic resonance imaging (fMRI) uses a powerful electromagnet to detect fluctuations in cerebral blood flow in different areas of the brain. Increased blood flow indicates increased activity, so this technology allows researchers to determine which areas of the brain are activated by different tasks and stimuli. Because a person must be able to tolerate the noise and close confinement of an MRI machine and must also be able to remain very still, most developmental fMRI studies have been done with children aged 6 years or older, often using practice sessions in a mock scanner to help children acclimate to the MRI environment. However, recent studies have used fMRI methods to investigate neural processes in sleeping infants as young as 2 days old. One such study demonstrated that the areas of the brain that are activated by spoken language from later infancy onward are also activated by speech in neonates (Perani et al., 2011).
Video: Interview with Charles Nelson
Other Techniques
Positron emission tomography (PET) measures brain activity by detecting the brain’s metabolic processes and has provided important information about brain development. However, because PET scans involve injecting radioactive material into the brain, this technique is used primarily for diagnostic purposes.
One of the newest methods to be used in developmental studies is near-infrared spectroscopy (NIRS), an optical imaging technique that measures neural activity by detecting metabolic changes that lead to differential absorption of infrared light in brain tissue. The infrared light is transmitted to the brain, and its absorption is detected by means of an optical-fiber skullcap or headband. Because NIRS is silent, noninvasive, and does not require rigid stabilization of the head, it is particularly promising for research with infants and young children. Thus far, a number of attempts to pinpoint infants’ brain activation in response to particular stimuli have shown mixed results, in part because the technology is still in its early stages (Aslin, 2012). One use of NIRS that has already proved successful is the monitoring of brain-oxygen levels in premature newborns. Another particularly interesting and successful application of NIRS involved a study of deaf children who had just received a cochlear implant, a surgically implanted electronic hearing device that we will discuss in Chapter 6. Because of concerns about the effects of its magnetic field, fMRI cannot be used to study brain processes in individuals with implants. NIRS was thus used to determine whether the auditory cortices of these children could respond to auditory stimulation. The NIRS showed that, rather remarkably, the auditory cortex of deaf children responded to sound within hours after the implant was activated, even though the cortex had never before been exposed to sound (Sevy et al., 2010).
After their “birth,” neurons begin the second developmental process, which involves migration to their ultimate destinations. Some neurons are pushed along passively by the newer cells formed after them, whereas others actively propel themselves toward their ultimate location.
spines formations on the dendrites of neurons that increase the dendrites’ capacity to form connections with other neurons
Once neurons reach their destination, cell growth and differentiation occur. Neurons first grow an axon and then a “bush” of dendrites (refer back to Figure 3.6). Thereafter, they take on the specific structural and functional characteristics of the different structures of the brain. Axons elongate as they grow toward specific targets, which, depending on the neuron in question, might be anything from another neuron in the brain to a bone in the big toe. The main change in dendrites is “arborization”—an enormous increase in the size and complexity of the dendritic “tree” that results from growth, branching, and the formation of spines on the branches. Arborization enormously increases the dendrites’ capacity to form connections with other neurons. In the cortex, the period of most intense growth and differentiation comes after birth.
111
112
myelination the formation of myelin (a fatty sheath) around the axons of neurons that speeds and increases information-processing abilities
The process of myelination, the formation of the insulating myelin sheath around some axons, begins in the brain before birth and continues into early adulthood. As noted earlier, a crucial function of myelin is to increase the speed of neural conduction. Myelination begins deep in the brain, beginning with the brainstem, and moves upward and outward into the cortex at a fairly steady rate throughout childhood and adolescence and into early adulthood (Lenroot & Giedd, 2006). The various cortical areas thus become myelinated at very different rates, possibly contributing to the different rates of development for various behaviors.
This pattern diverges in interesting ways from the pattern found in our primate relative, the chimpanzee. Initially, white matter develops even more slowly in the prefrontal lobes of infant and juvenile chimpanzees than it does in young humans, suggesting one possible mechanism whereby evolutionary pressure has improved human brain function relative to that of other primates (Sakai et al., 2011). Interestingly, however, the chimpanzee shows a mature pattern of myelination at the point of sexual maturity, far earlier than that observed in humans (D. J. Miller et al., 2012). The reasons for the extended period of human myelination remain unknown, and might be both positive (facilitating improvements in executive functions after sexual maturity) and negative (making the human brain more vulnerable to disorders related to myelination).
Synaptogenesis
synaptogenesis the process by which neurons form synapses with other neurons, resulting in trillions of connections
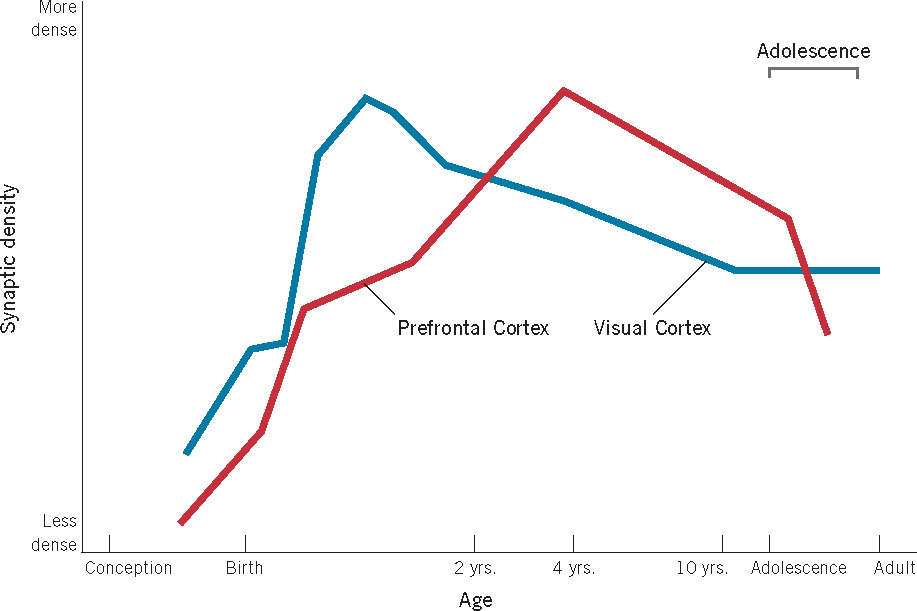
One result of the extraordinary growth of axonal and dendritic fibers is a wildly exuberant generation of neuronal connections. In a process called synaptogenesis, each neuron forms synapses with thousands of others, resulting in the formation of the trillions of connections referred to earlier. Figure 3.8 shows the time course of synaptogenesis in the cortex. As you can see, it begins prenatally and proceeds very rapidly both before birth and for some time afterward. Note that both the timing and rate of synapse production vary for different cortical areas; synapse generation is complete much earlier in the visual cortex, for example, than in the frontal area. As with myelination, the differential timing of synapse generation across areas of the brain likely contributes to the developmental timing of the onset of various abilities and behaviors.
113
Synapse Elimination
The explosive generation of neurons and synapses during synaptogenesis, which is largely under genetic control, results in a huge surplus—many more neural connections than any one brain can use. This overabundance of synapses includes an excess of connections between different parts of the brain: for instance, many neurons in what will become the auditory cortex are linked with those in the visual area, and both of these areas are overly connected to neurons involved in taste and smell. As a consequence of this hyperconnectivity, newborns may experience synesthesia—the blending of different types of sensory input (Maurer & Mondloch, 2004). In the case of the extra connections between auditory and visual cortex, for example, auditory stimulation may produce a visual experience, with the infant’s perceiving a sound as being of a particular color.
synaptic pruning the normal developmental process through which synapses that are rarely activated are eliminated
We now come to what is one of the most remarkable facts about the development of the human brain. Approximately 40% of this great synaptic superfluity gets eliminated in a developmental process known as synaptic pruning. As you learned in the previous chapter, cell death is a normal part of development, and nowhere is that more evident than in the systematic pruning of excess synapses that continues for years after birth. This pruning occurs at different times in different areas of the brain (P. R. Huttenlocher & Dabholkar, 1997). You can see from Figure 3.8 that the elimination of synapses in the visual cortex begins near the end of the first year of life and continues until roughly 10 years of age, whereas synapse elimination in the prefrontal area shows a slower time course. During peak pruning periods, as many as 100,000 synapses may be eliminated per second (Kolb, 1995)!
Not known until fairly recently is the fact that the brain undergoes explosive changes during adolescence, including a wave of overproduction and pruning akin to that in the first years of life (Giedd et al., 1999; Gogtay et al., 2004). Although the amount of white matter in the cortex shows a steady increase from childhood well into adulthood, the amount of gray matter increases dramatically starting around 11 or 12 years of age. The increase in gray matter proceeds rapidly, peaks around puberty, and then begins to decline as some of it is replaced by white matter (see Figure 3.9). The last area of the cortex to mature is the dorsolateral prefrontal cortex, which is vital for regulating attention, controlling impulses, foreseeing consequences, setting priorities, and other executive functions. It does not reach adult dimensions until after the age of 20.
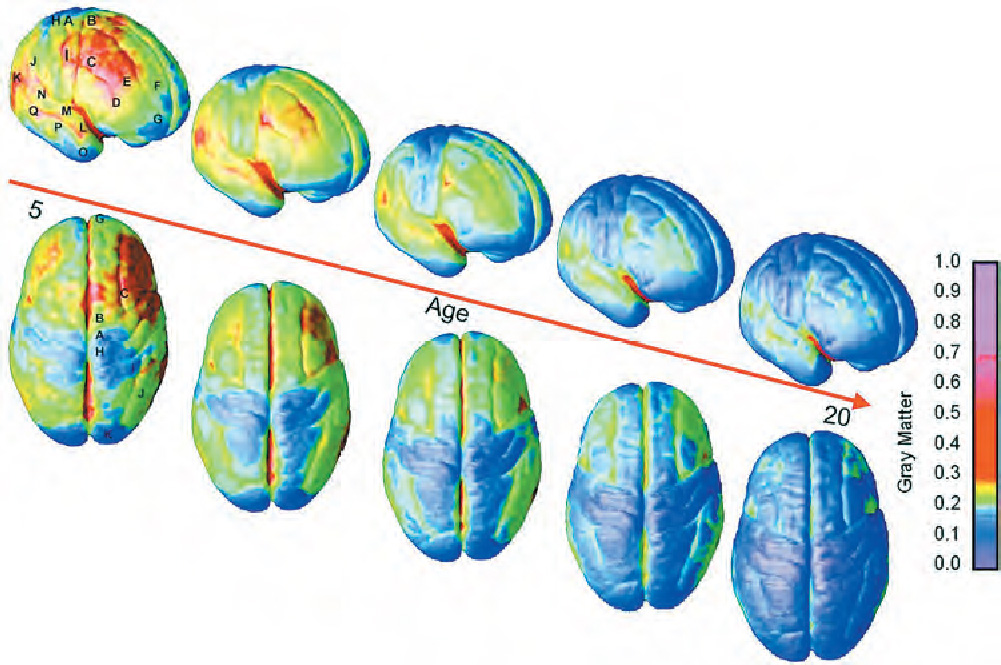
You will notice that Figure 3.8 does not show a second proliferation and reduction of synaptic density in adolescence. One of the reasons for this is that the figure is based on cross-sectional research in which the brains of individuals of different ages were examined only at autopsy. The new data showing substantial development in the adolescent brain come from longitudinal studies in which the same individuals’ brains were scanned repeatedly over several years. The dramatic changes that appear in individuals were not evident when the brains of separate groups of different ages were studied. (See the discussion of developmental research designs in Chapter 1.)
114
The Importance of Experience
What factors determine which of the brain’s excess synapses will be pruned and which maintained? Experience plays a central role in what is essentially a case of “use it or lose it.” In a competitive process that has been dubbed “neural Darwinism” (Edelman, 1987), those synapses that are frequently activated are selectively preserved (Changeux & Danchin, 1976). The more often a synapse is activated, the stronger the connection becomes between the neurons involved: in short, neurons that fire together wire together (Hebb, 1949). Conversely, when a synapse is rarely active, it is likely to disappear: the axon of one neuron withdraws and the dendritic spine of the other is “pruned away.”
plasticity the capacity of the brain to be affected by experience
The obvious question now is: Why does the human brain—the product of millions of years of evolution—take such a devious developmental path, producing a huge excess of synapses, only to get rid of a substantial proportion of them? The answer appears to be evolutionary economy. The capacity of the brain to be molded or changed by experience, referred to as plasticity, means that less information needs to be encoded in the genes. This economizing may, in fact, be a necessity: the number of genes involved in the formation and functioning of the nervous system is enough to specify only a very small fraction of the normal complement of neurons and neural connections. In addition, if brain structures were entirely hard-wired, organisms would be unable to adapt to their postnatal environment. To complete the final wiring of the brain, nurture joins forces with nature.
The collaboration between nature and nurture in building the brain occurs differently for two kinds of plasticity. One kind involves the general experiences that almost all infants have just by virtue of being human. The second kind involves specific, idiosyncratic experiences that children have as a result of their particular life circumstances—such as growing up in the United States or in the Amazon rain forest, experiencing frequent cuddling or abuse, being an only child or one of many siblings, and so on.
115
Experience-Expectant Processes
experience-expectant plasticity the process through which the normal wiring of the brain occurs in part as a result of experiences that every human who inhabits any reasonably normal environment will have
William Greenough refers to the role of general human experience in shaping brain development as experience-expectant plasticity. According to this view, the normal wiring of the brain is in part a result of the kinds of general experiences that have been present throughout human evolution, experiences that every human with an intact sensory-motor system who inhabits a reasonably normal environment will have: patterned visual stimulation, voices and other sounds, movement and manipulation, and so forth (Greenough & Black, 1992). Consequently, the brain can “expect” input from these reliable sources to fine-tune its circuitry; synapses that are frequently activated will be strengthened and stabilized and those that are rarely activated will be “pruned.” Thus, our experience of the external world plays a fundamental role in shaping the most basic aspects of the structure of our brain.
One fundamental benefit of experience-expectant plasticity is that, because experience helps shape the brain, fewer genes need to be dedicated to normal development. Another is that the brain is better able to recover from injury to certain areas, because other brain areas can take over the function that would have been performed by the damaged area. The younger the brain when damaged, the more likely recovery is.
The downside of experience-expectant plasticity is that it is accompanied by vulnerability. If for some reason the experience that the developing brain is “expecting” for fine-tuning its circuits does not occur, whether because of inadequate stimulation or impaired sensory receptors, development may be compromised. A good example of this vulnerability comes from children who are born with cataracts that obscure their vision. The longer a cataract remains in place after birth, the more impaired the child’s visual acuity will be once it is removed. Dramatic improvement typically follows early removal, although some aspects of visual processing (especially of faces) remain affected even into adulthood (de Heering & Maurer, 2012; Maurer, Mondloch, & Lewis, 2007). Presumably, the lasting deficits of late cataract removal occur because synapses that would normally have been activated by visual stimulation after birth were pruned because of the lack of that stimulation.
When an expected form of sensory experience is absent, what happens to areas of the brain that normally would have become specialized as a result of that experience? A wealth of data from animals indicates that such areas can become at least partially reorganized to serve some other function. Evidence of such plasticity and reorganization in humans comes from studies of congenitally deaf adults who, as children, had learned American Sign Language, a full-fledged, visually based, language (Bavelier, Dye, & Hauser, 2006; Bavelier & Neville, 2002). Deaf individuals rely heavily on peripheral vision for language processing; they typically look into the eyes of a person who is signing to them, while using their peripheral vision to monitor the hand and arm motions of the signer. ERP recordings of brain activity (see Box 3.3) showed that deaf individuals’ responses to peripheral visual stimuli are several times stronger than those of hearing people. In addition, their responses are distributed differently across brain regions. Thus, because of the lack of auditory experience, brain systems that would normally be involved in hearing and in spoken-language processing become organized to process visual information instead.
Similar evidence of early brain reorganization comes from research with blind adults. When tested for their ability to discriminate changes in musical pitch, adults who were born blind or became blind quite early performed much better than those who had become blind later in life (Gougoux et al., 2004). Presumably, connections between visual and auditory cortex were preserved in individuals with early-onset blindness, giving them extra “brain power” to apply to the auditory task. Consistent with this idea, brain-imaging research suggests that parts of the visual cortex contribute to superior sound localization ability in adults with early-onset blindness. A related result is that congenitally blind individuals show activation in the “visual” cortex both when reading Braille (Sadato et al., 1998) and when processing spoken language (Bedny et al., 2011).
116
Sensitive periods As suggested by the foregoing examples, a key element in experience-expectant plasticity is timing. There are a few sensitive periods when the human brain is especially sensitive to particular kinds of external stimuli. It is as though a time window were temporarily opened, inviting environmental input to help organize the brain. Gradually, the window closes. The neural organization that occurs (or does not occur) during sensitive periods is typically irreversible.
As discussed in Chapter 1, the extreme deprivation that the Romanian orphans suffered early in life, when children normally experience a wealth of social and other environmental stimulation, is considered by some to be an example of a sensitive-period effect. Some investigators speculate that adolescence, during which rapid changes are occurring in the brain, may be another sensitive period for various aspects of development. Yet another sensitive period, for language learning, will be discussed in Chapter 6.
Experience-Dependent Processes
experience-dependent plasticity the process through which neural connections are created and reorganized throughout life as a function of an individual’s experiences
The brain is also sculpted by idiosyncratic experience through what Greenough calls experience-dependent plasticity. Neural connections are created and reorganized constantly, throughout life, as a function of an individual’s experiences. (If you remember anything of what you have been reading in this chapter, it’s because you have formed new neural connections.)

ENVIRONMENT IMAGES/UIG/GETTYIMAGES
Much of the research on experience-dependent plasticity has been focused on nonhuman animals, whose environments can be readily manipulated. One such method has involved comparisons between animals reared in complex environments full of objects to explore and use versus animals reared in bare laboratory cages. The brains of rats (and cats and monkeys) that grow up in a complex environment have more dendritic spines on their cortical neurons, more synapses per neuron, and more synapses overall, as well as a generally thicker cortex and more of the supportive tissues (such as blood vessels and glial cells) that maximize neuronal and synaptic function. All this extra hardware seems to have a payoff: rats (and other animals) reared in a complex environment (which is more akin to their natural environment) perform better in a variety of learning tasks than do their counterparts raised in bare cages (e.g., Sale, Berardi, & Maffei, 2009).

DAVID RAMOS/GETTY IMAGES
Highly specific effects of experience on brain structure also occur. For example, rats that are trained to use just one forelimb to get a food reward have increased dendritic material in the particular area of the motor cortex that controls the movement of the trained limb (Greenough, Larson, & Withers, 1985). In humans, research on musicians has revealed that, compared with a control group, violinists and cellists had increased cortical representation of the fingers of the left hand (Elbert et al., 1995). In other words, after years of practice, more cortical cells were devoted to receiving input from and controlling the fingers that manipulate the strings of the instruments. Similarly, in skilled Braille readers, the cortical representation of the left hand—which is used to read Braille text—is enlarged (Pascual-Leone et al., 1993).
117
Effects of specific experience are also evident in fMRI studies of individuals with dyslexia, a severe reading problem in people with normal intelligence and schooling (see Chapter 8). One example involves a remedial reading program in which 2nd- and 3rd-graders with dyslexia received training in recognizing the correspondence between speech sounds and letters (Blachman et al., 2004). After the training, not only did the children show marked improvement in their reading ability, but fMRI imaging revealed increased activity in their left-brain areas that was similar to the activity in the brains of good readers. The specific effects of one’s reading experience also show up in the fact that reading Chinese characters recruits distinctly different brain networks than those involved in reading an alphabetic script (such as English).
Brain Damage and Recovery
As noted previously, because of its plasticity (especially early in life), the brain can become rewired—at least to some degree—after suffering damage. Children who suffer from brain damage thus have a better chance of recovering lost function than do adults who suffer similar damage. The strongest evidence for this comes from young children who suffer damage to the language area of the cortex and who generally recover most, if not all, of their language functions. This is because after the damage has occurred, other areas of the immature brain can take over language functions. As a result, language is largely spared, though specific linguistic impairments may remain (e.g., Zevin, Datta, & Skipper, 2012).
In contrast, adults who sustain the same type of brain damage undergo no such reorganization of language functions and may have a permanent loss in the ability to comprehend or produce speech. Greater recovery from early brain injury has also been observed for functions other than language. For example, producing appropriate facial expressions is more difficult for adults who had damage to the frontal area of the cortex during adulthood than for adults whose frontal lobe injury occurred in childhood (Kolb, 1995).
It is not always true, however, that the chance of recovery from early brain injury is greater than it is for later injury. Likelihood of recovery depends on how extensive the damage is and what aspect of brain development is occurring at the time of the damage. Consider, for example, the offspring of Japanese women who, while pregnant, were exposed to massive levels of radiation from the atomic bombs dropped on Hiroshima and Nagasaki in 1945. The rate of intellectual impairment was much higher for surviving children whose exposure had occurred very early in prenatal development, during the time of rapid neurogenesis and migration of neurons (Otake & Schull, 1984). Similarly, brain injury during early childhood generally results in more severe cognitive impairment in IQ than does later comparable injury (V. Anderson et al., 2012).
118
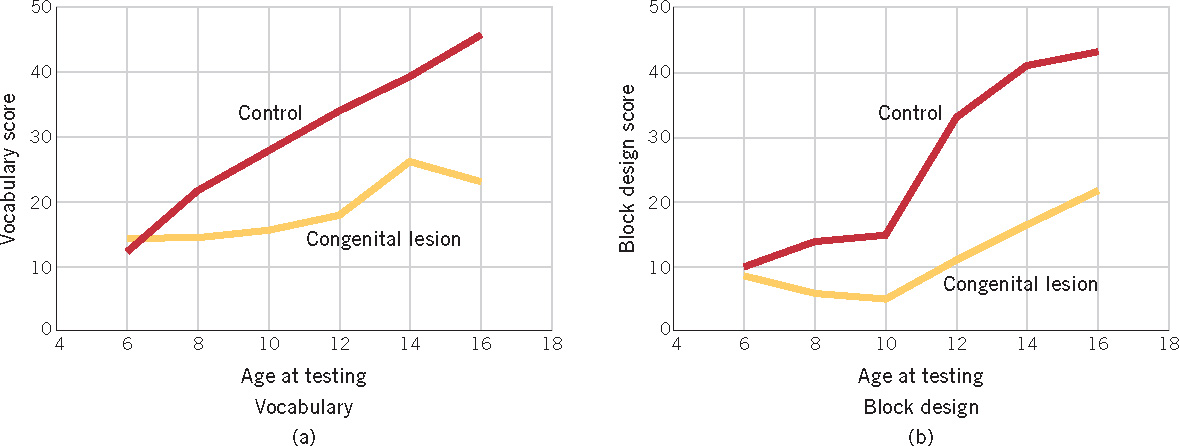
Furthermore, even when children appear to have made a full recovery from an early brain injury, deficits may emerge later. This was demonstrated in a cross-sectional study that compared cognitive performance in a group of children who had been born with cerebral damage and a control group of children with no brain damage (Banich et al., 1990). As Figure 3.10 shows, the children with brain damage did not differ from the control group in their performance on two subscales of an IQ test at 6 years of age. However, as the normal children’s performance improved with age, the brain-damaged children’s performance fell progressively behind. The same pattern of results—decline in IQ over age for children with brain damage—was also demonstrated in a longitudinal study, in which children who had sustained early damage were tested before and after age 7 (S. C. Levine et al., 2005). These results illustrate the difficulty of predicting the development of children with cerebral injuries: behavior that appears normal early in development may deteriorate.
On the basis of these various aspects of plasticity, we can generalize that the worst time to suffer brain damage is very early, during prenatal development and the first year after birth, when neurogenesis is occurring and basic brain structures are being formed. Damage at this point may have cascading effects on subsequent aspects of brain development, with potentially wide-ranging negative effects. In contrast, when brain damage is sustained in early childhood—that is, when synapse generation and pruning are occurring and plasticity is highest—the chances for the brain’s rewiring itself and recovering lost function are best.
review:
Nature and nurture cooperate in the construction of the human brain. Some important brain structures include the neurons, which communicate with one another at synapses; the cortex, in which different functions are localized in different areas; and the cerebral hemispheres, which are specialized for different kinds of processing. The processes involved in the development of the brain include neurogenesis and synaptogenesis, followed by the systematic elimination of some synapses and the preservation of others as a function of experience.
Two forms of plasticity contribute to the development of behavior. As a result of experience-expectant plasticity, the brain is shaped by experiences that are available to every typically developing individual in interaction with every species-typical environment. Through experience-dependent plasticity, the brain is also structured by an individual’s idiosyncratic life experiences. Because of the importance of experience in brain development, sensitive periods exist during which specific experience must be present for normal development. Timing is also a crucial factor in the ultimate impact of brain damage.
119