Chapter 15
The highly integrated biochemical reactions that take place inside the cell
Anabolism is the set of biochemical reactions that use energy to build new molecules and, ultimately, new cells. Catabolism is the set of biochemical reactions that extract energy from fuel sources or breakdown biomolecules.
Page C15You reply that vandalism is disrespectful and expensive. Part of your tuition money will now have to pay to remove the vandalism. Plus, the fool should know that Gibbs free energy is at a minimum when a system is in equilibrium.
(1) Cellular movements and the performance of mechanical work; (2) active transport; (3) biosynthetic reactions
Complete the interactive matching exercise to see answers.
Charge repulsion, resonance stabilization, increase in entropy, and stabilization by hydration
Having only one nucleotide function as the energy currency of the cell enables the cell to monitor its energy status.
These divalent ions bind to the negatively charged oxygen atoms found on the phosphoryl groups and help stabilize the charges on ATP.
Increasing the concentration of ATP or decreasing the concentration of cellular ADP or Pi (by rapid removal by other reactions, for instance) would make the reaction more exergonic. Likewise, altering the Mg2+ concentration could raise or lower the ΔG of the reaction (problems 25 and 30).
Reactions in parts a and c, to the left; reactions in parts b and d, to the right
None whatsoever
(a) ΔG°′ = 31.4 kJ mol−1 (7.5 kcal mol−1) and
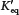 = 3.06 × 10−6; (b) 3.28 × 104
= 3.06 × 10−6; (b) 3.28 × 104ΔG°′ = 7.1 kJ mol−1 (1.7 kcal mol−1). The equilibrium ratio is 17.8.
Acetate + CoA + H+ goes to acetyl CoA + H2O, ΔG°′ = +31.4 kJ mol−1 (+7.5 kcal mol−1). ATP hydrolysis to AMP, ΔG°′ = 45.6 kJ mol−1 (−10.9 kcal mol−1). Overall reaction, ΔG°′ = −14.2 kJ mol−1 (−3.4 kcal mol−1).
With pyrophosphate hydrolysis, ΔG°′ = −33.5 kJ mol−1 (−8.0 kcal mol−1). Pyrophosphate hydrolysis dramatically increases the exergonicity of the reaction.
The free-
energy changes of the individual steps in a pathway are summed to determine the overall free- energy change of the entire pathway. Consequently, a reaction with a positive free- energy value can be powered to take place if coupled to a sufficiently exergonic reaction. An ADP unit
Higher organisms cannot make vitamins, and thus are dependent on obtaining them from other organisms.
NADH and FADH2 are electron carriers for catabolism; NADPH is the carrier for anabolism.
The electrons of the C=O bond cannot form resonance structures with the C—
S bond that are as stable as those that they can form with the C— O bond. Thus, the thioester is not stabilized by resonance to the same degree as is an oxygen ester. (1) Control of the amount of enzymes; (2) control of enzyme activity; (3) control of the availability of substrates
Recall that ΔG = ΔG°′ + RT ln [products]/[reactants]. Altering the ratio of products to reactants will cause ΔG to vary. In glycolysis, the concentrations of the components of the pathway result in a value of ΔG greater than that of ΔG°′.
A, Ethanol; B, lactate; C, succinate; D, isocitrate; E, malate
Unless the ingested food is converted into molecules capable of being absorbed by the intestine, no energy can ever be extracted by the body.
Although the reaction is thermodynamically favorable, the reactants are kinetically stable because of the large activation energy. Enzymes lower the activation energy so that the reaction takes place on time scales required by the cell.
For an acid AH,
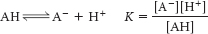
The pKa is defined as pKa = −log10 K.
ΔG°′ is the standard free-
energy change at pH 7. Thus, ΔG°′ = −RT ln K = −2.303 RT log10 K = +2.303 RT pKa. ΔG°′ = 27.32 kJ mol−1 (6.53 kcal mol−1)
The activated form of sulfate in most organisms is 3′-phosphoadenosine-
5′-phosphosulfate. As the Mg2+ concentration falls, the ΔG of hydrolysis rises. Note that pMg is a logarithmic plot, and so each number on the x axis represents a 10-
fold change in [Mg2+]. Mg2+ binds to the phosphoryl groups of ATP and helps to mitigate charge repulsion. As the [Mg2+] falls, there is less charge stabilization of ATP, leading to greater charge repulsion and an increase in ΔG on hydrolysis.
Arginine phosphate in invertebrate muscle, like creatine phosphate in vertebrate muscle, serves as a reservoir of high-
potential phosphoryl groups. Arginine phosphate maintains a high level of ATP in muscular exertion. The rationale behind creatine supplementation is that it would be converted into creatine phosphate and thus serve as a rapid means of replenishing ATP after muscle contraction.
If creatine supplementation were beneficial, it would affect activities that depend on short bursts of activity; any sustained activity would require ATP generation by fuel metabolism, which, as Figure 15.8 shows, requires more time.
Under standard conditions, ΔG°′ = −RT ln [products]/[reactants]. Substituting 23.8 kJ mol−1 (5.7 kcal mol−1) for ΔG°′ and solving for [products]/[reactants] yields 7 × 10−5. In other words, the forward reaction does not take place to a significant extent. Under intracellular conditions, ΔG is −1.3 kJ mol−1 (−0.3 kcal mol−1). Using the equation ΔG = ΔG°′ + RT ln [product]/[reactants] and solving for [products]/[reactants] gives a ratio of 3.7 × 10−5. Thus, a reaction that is endergonic under standard conditions can be converted into an exergonic reaction by maintaining the [products]/[reactants] ratio below the equilibrium value. This conversion is usually attained by using the products in another coupled reaction as soon as they are formed.
Under standard conditions,
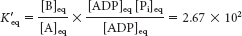
At equilibrium, the ratio of [B] to [A] is given by
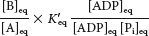
The ATP-
generating system of cells maintains the [ATP]/[ADP][Pi] ratio at a high level, typically about 500 M−1. For this ratio, 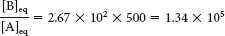
This equilibrium ratio is strikingly different from the value of 1.15 × 10−3 for the reaction A → B in the absence of ATP hydrolysis. In other words, coupling the hydrolysis of ATP with the conversion of A into B has changed the equilibrium ratio of B to A by a factor of about 108.
Liver: −45.2 kJ mol−1 (−10.8 kcal mol−1); muscle: −48.1 kJ mol−1 (−11.5 kcal mol−1); brain: −48.5 kJ mol−1 (−11.6 kcal mol−1). The free energy of ATP hydrolysis is most negative in the brain.