Chapter 28
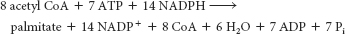
The first reaction
Oxaloacetate + NADH + H+ ⇌ malate + NAD+
is catalyzed by cytoplasmic malate dehydrogenase. The next reaction is catalyzed by malic enzyme:
Malate + NADP+ → pyruvate + CO2 + NADPH
Finally, oxaloacetate is regenerated from pyruvate by pyruvate carboxylase:
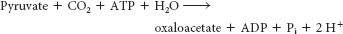
The sum of the reactions is
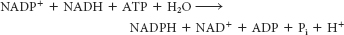
The formation of malonyl CoA from acetyl CoA by acetyl CoA carboxylase 1
Complete the interactive matching exercise to see answers.
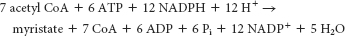
Six acetyl CoA units are required. One acetyl CoA unit is used directly to become the two carbon atoms farthest from the acid end (the ω end). The other five units must be converted into malonyl CoA. The synthesis of each malonyl CoA molecule costs a molecule of ATP; so 5 molecules of ATP are required. Each round of elongation requires 2 molecules of NADPH, one to reduce the keto group to an alcohol and one to reduce the double bond. As a result, 10 molecules of NADPH are required. Therefore, 5 molecules of ATP and 10 molecules of NADPH are required to synthesize lauric acid.
e, b, d, a, c
Such a mutation would inhibit fatty acid synthesis because the enzyme cleaves cytoplasmic citrate to yield acetyl CoA for fatty acid synthesis.
False. Biotin is required for acetyl CoA carboxylase activity.
True
False. ATP is required to synthesize malonyl CoA.
True
True
False. Fatty acid synthase is a dimer.
True
False. Acetyl CoA carboxylase 1 is stimulated by citrate, which is cleaved to yield its substrate acetyl CoA.
Fatty acids with odd numbers of carbon atoms are synthesized starting with propionyl ACP (instead of acetyl ACP), which is formed from propionyl CoA by acetyl transacylase.
The only acetyl CoA used directly, not in the form of malonyl CoA, provides the two carbon atoms at the ω end of the fatty acid chain. Because palmitic acid is a C16 fatty acid, acetyl CoA will have provided carbons 15 and 16.
Page C30HCO3− is attached to acetyl CoA to form malonyl CoA. When malonyl CoA condenses with acetyl CoA to form the four-
carbon ketoacyl CoA, the HCO3− is lost as CO2. Bicarbonate is required for the synthesis of malonyl CoA from acetyl CoA by acetyl CoA carboxylase.
C-
1 is more radioactive. Decarboxylation drives the condensation of malonyl ACP and acetyl ACP. In contrast, the condensation of two molecules of acetyl ACP is energetically unfavorable. In gluconeogenesis, decarboxylation drives the formation of phosphoenolpyruvate from oxaloacetate.
The mutant enzyme will be persistently active because it cannot be inhibited by phosphorylation. Fatty acid synthesis will be abnormally active. Such a mutation might lead to obesity.
The probability of synthesizing an error-
free polypeptide chain decreases as the length of the chain increases. A single mistake can make the entire polypeptide ineffective. In contrast, a defective subunit can be spurned in the formation of a noncovalent multienzyme complex; the good subunits are not wasted. When glycogen stores are filled, the excess carbohydrates are metabolized to acetyl CoA, which is then converted into fats. Human beings cannot convert fats into carbohydrates, but they can certainly convert carbohydrates into fats.
Cytoplasmic palmitoyl CoA inhibits the translocase that shuttles citrate from mitochondria to the cytoplasm, thereby preventing fatty acid synthesis. Mitochondrial malonyl CoA inhibits carnitine acyltransferase I, preventing the entry of fatty acids into mitochondria, thereby inhibiting fatty acid degradation in times of plenty.
With a diet rich in raw eggs, avidin will inhibit fatty acid synthesis by reducing the amount of biotin required by acetyl CoA carboxylase. Cooking the eggs will denature avidin, and so it will no longer bind biotin.
Phosphofructokinase controls the flux down the glycolytic pathway. Glycolysis functions to generate ATP or building blocks for biosynthesis, depending on the tissue. The presence of citrate in the cytoplasm indicates that those needs are met, and there is no need to metabolize glucose.
Oxidation in mitochondria; synthesis in the cytoplasm
Coenzyme A in oxidation; acyl carrier protein in synthesis
FAD and NAD+ in oxidation; NADPH in synthesis
l isomer of 3-
hydroxyacyl CoA in oxidation; d isomer in synthesis From carboxyl to methyl in oxidation; from methyl to carboxyl in synthesis
The enzymes of fatty acid synthesis, but not those of oxidation, are organized in a multienzyme complex.
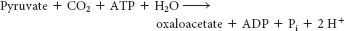
In gluconeogenesis, the oxaloacetate is converted into phosphoenolpyruvate by PEP carboxykinase at the expense of a molecule of GTP.
The product of the reaction catalyzed by acetyl CoA carboxylase 2, malonyl CoA, inhibits the import of fatty acids into mitochondria by inhibiting carnitine acyltransferase I.
Citrate is an allosteric activator of acetyl CoA carboxylase 1, causing the enzyme to form active filaments. Palmitoyl CoA will inhibit the enzyme.
Soraphen A inhibits fatty acid synthesis in a dose-
dependent manner. Fatty acid oxidation is increased in the presence of soraphen A.
Recall that acetyl carboxylase 2 synthesizes malonyl CoA to inhibit the transport of fatty acids into the mitochondria, thereby preventing fatty acid oxidation. Soraphen A apparently inhibits both forms of the carboxylase.
Phospholipid synthesis was inhibited in a dose-
dependent manner. Phospholipids are required for membrane synthesis.
Soraphen A inhibits cell proliferation, especially at higher concentrations.
All of the labeled carbon atoms will be retained. Because you need 8 acetyl CoA molecules and only 1 carbon atom is labeled in the acetyl group, you will have 8 labeled carbon atoms. The only acetyl CoA used directly will retain 3 tritium atoms. The 7 acetyl CoA molecules used to make malonyl CoA will lose 1 tritium atom on addition of the CO2 and another tritium atom at the dehydration step. Each of the 7 malonyl CoA molecules will retain 1 tritium atom. Therefore, the total retained tritium is 10 atoms. The ratio of tritium to carbon is 1.25.
The high concentration of NADH inhibits gluconeogenesis by preventing the oxidation of lactate to pyruvate while causing the reverse reaction to predominate, leading to lactic acidosis and hypoglycemia. The NADH glut also inhibits fatty acid oxidation and the citric acid cycle because the drinker’s NADH needs are met by ethanol metabolism. The excess NADH signals that conditions are right for fatty acid synthesis, which leads to the development of “fatty liver.”