12.5 A Major Role of Membrane Proteins Is to Function as Transporters
Transporter proteins are a specific class of pump or channel that facilitates the movement of molecules across a membrane. Each cell type expresses a specific set of transporters in its plasma membrane. The set of transporters expressed is crucial because these transporters largely determine the ionic composition inside a cell and the compounds that can be taken up from the cell’s environment. In some senses, the array of transporters expressed by a cell determines its characteristics because a cell can execute only those biochemical reactions for which it has taken up the substrates.
Two factors determine whether a small molecule will cross a membrane: (1) the concentration gradient of the molecule across the membrane and (2) the molecule’s solubility in the hydrophobic environment of the membrane. In accord with the Second Law of Thermodynamics, molecules spontaneously move from a region of higher concentration to one of lower concentration. For many molecules, the cell membrane is an obstacle to this movement, but, as discussed earlier, some molecules can pass through the membrane because they dissolve in the lipid bilayer. Such molecules are called lipophilic molecules. The steroid hormones provide a physiological example. These cholesterol relatives can pass through a membrane in their path of movement in a process called simple diffusion.
Matters become more complicated when the molecule is highly polar. For example, sodium ions are present at 143 mM outside a typical cell and at 14 mM inside the cell, yet sodium does not freely enter the cell, because the charged ion cannot pass through the hydrophobic membrane interior. In some circumstances, such as in a nerve impulse, sodium ions must enter the cell. How are these ions able to cross the hydrophobic membrane interior? Sodium ions pass through specific channels in the hydrophobic barrier—
How is the sodium gradient established in the first place? In this case, sodium must move, or be pumped, against a concentration gradient. Because moving the ion from a low concentration to a higher concentration results in a decrease in entropy, it requires an input of free energy. Protein pumps embedded in the membrane are capable of using an energy source to move the molecule up a concentration gradient in a process called active transport.
The Na+–K+ ATPase Is an Important Pump in Many Cells
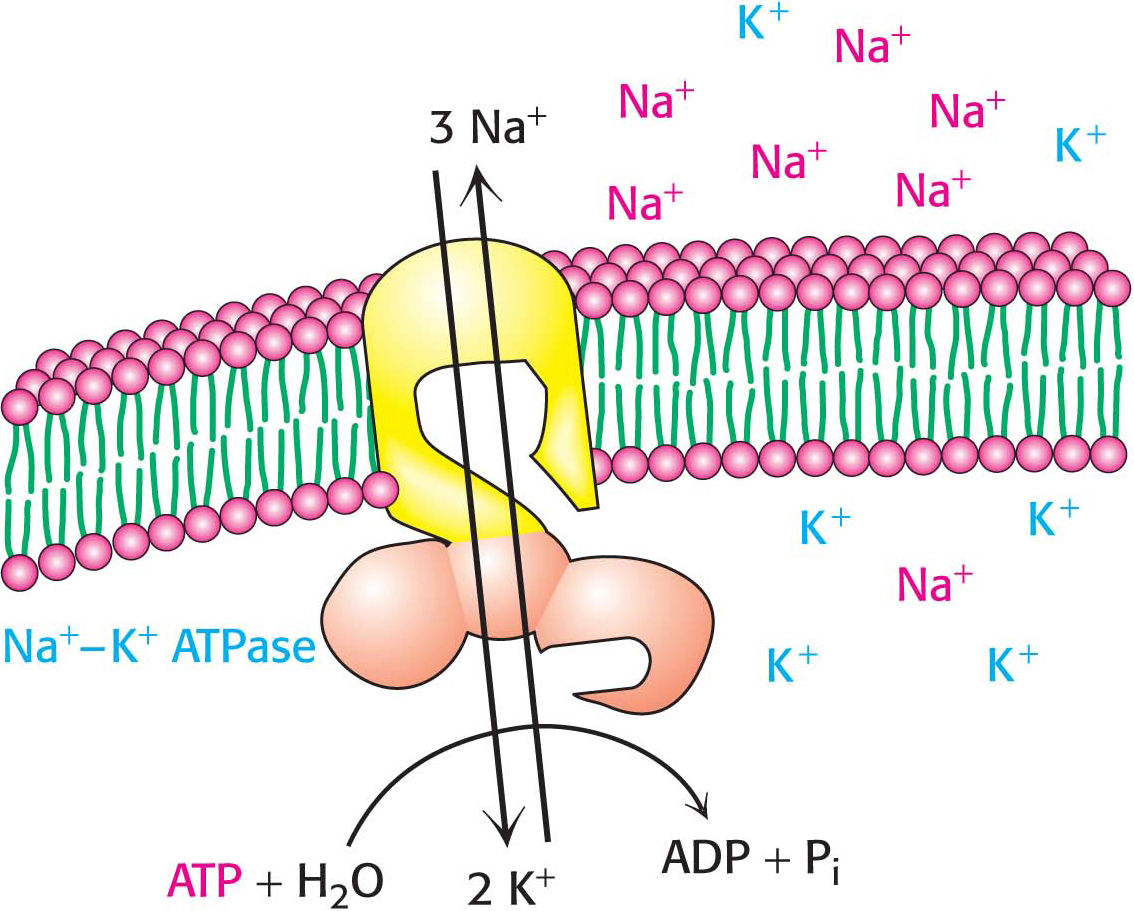
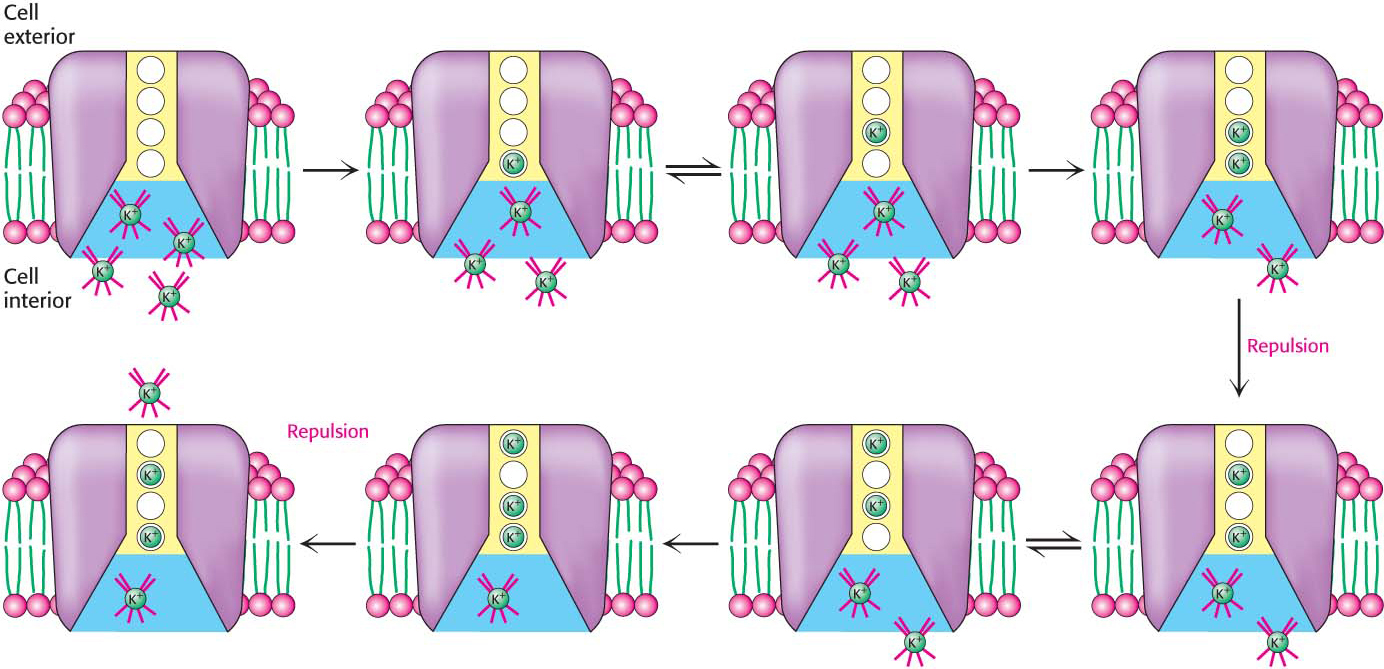
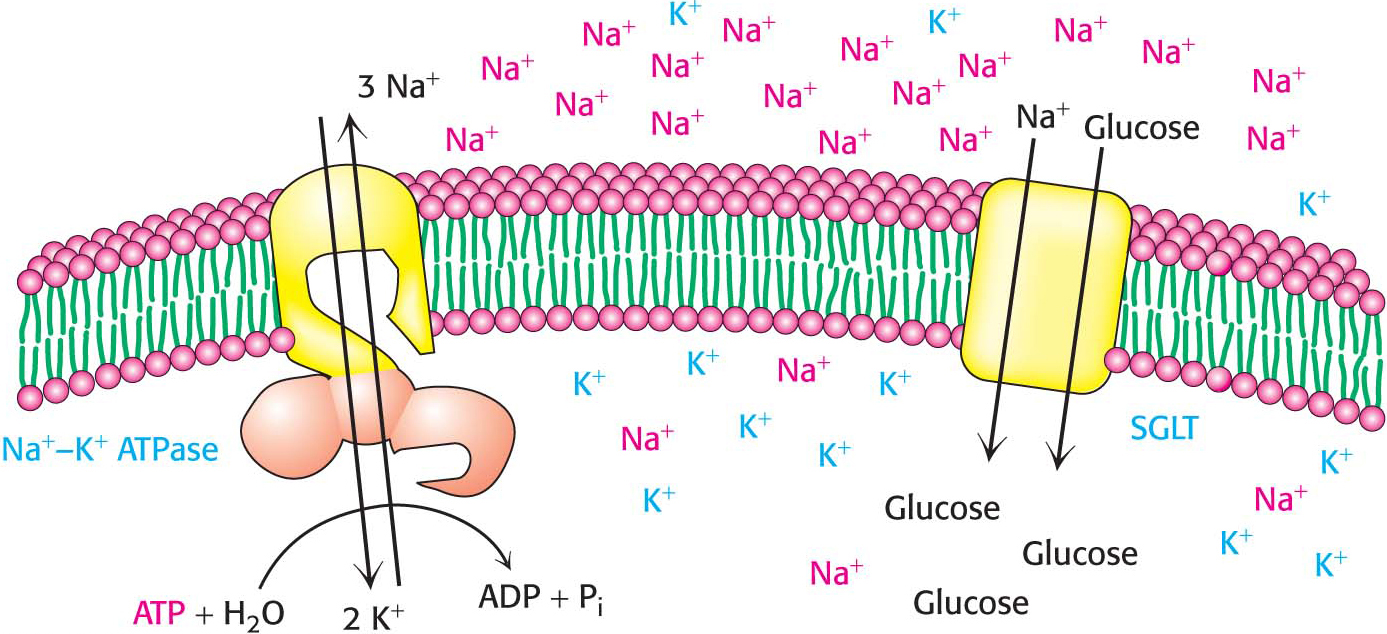
Cells must control their intracellular salt concentrations to prevent unfavorable interactions with high concentrations of ions and to facilitate specific processes such as a nerve impulse. In particular, most animal cells contain a high concentration of K+ and a low concentration of Na+ relative to the extracellular fluid. These ionic gradients are generated by a specific transport system, an enzyme called the Na+–K+ pump or the Na+–K+ ATPase. The hydrolysis of ATP by the pump provides the energy needed for the active transport of three Na+ ions out of the cell and two K+ ions into the cell, generating the gradients (Figure 12.16). In other words, the Na+–K+ ATPase is an ATP-
The active transport of Na+ and K+ is of great physiological significance. Indeed, more than a third of the ATP consumed by a resting animal is used to pump these ions. The Na+–K+ gradient in animal cells controls cell volume, renders neurons and muscle cells electrically excitable, and drives the active transport of sugars and amino acids. This third phenomenon is called secondary active transport because the sodium gradient generated by the Na+–K+ ATPase (the primary instance of active transport) can be used to power active transport of other molecules (the secondary instance of active transport) when the sodium flows down its gradient.
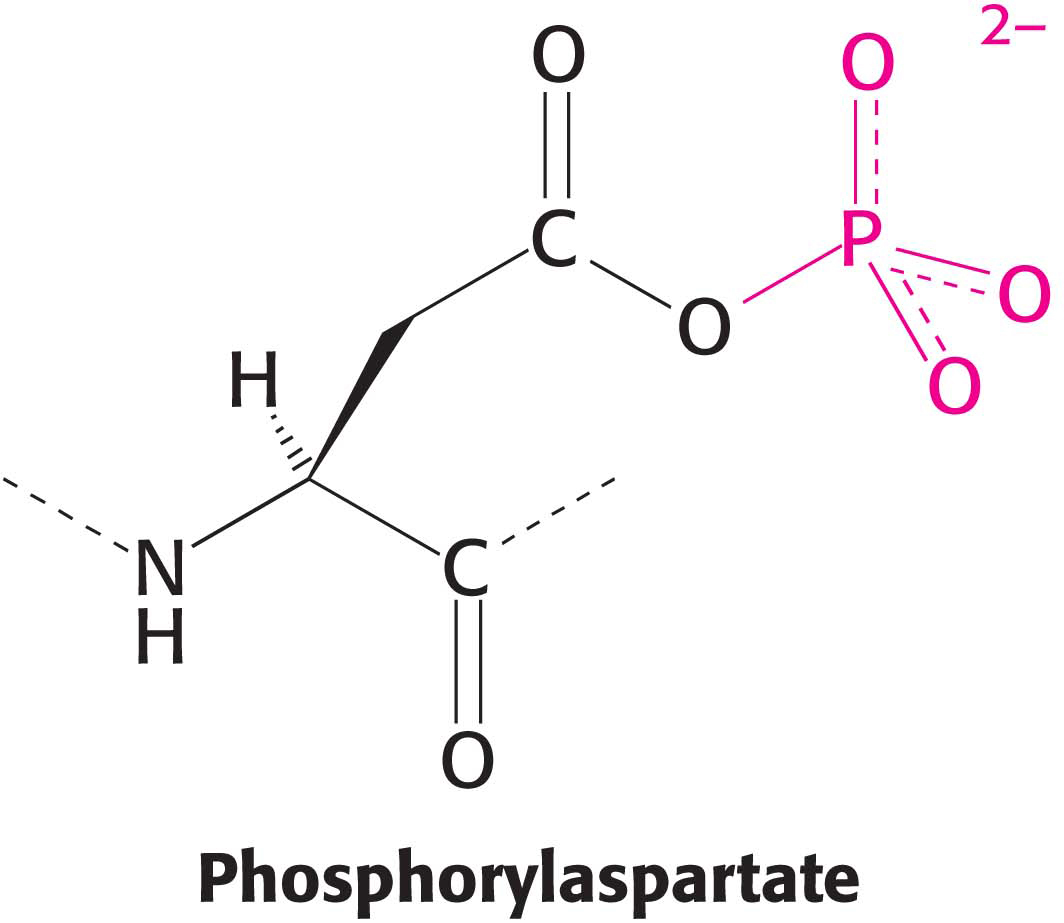
The pump is called the Na+–K+ ATPase because the hydrolysis of ATP takes place only when Na+ and K+ are present. The Na+–K+ ATPase is referred to as a P-
 CLINICAL INSIGHT
CLINICAL INSIGHTMultidrug Resistance Highlights a Family of Membrane Pumps with ATP-Binding Domains
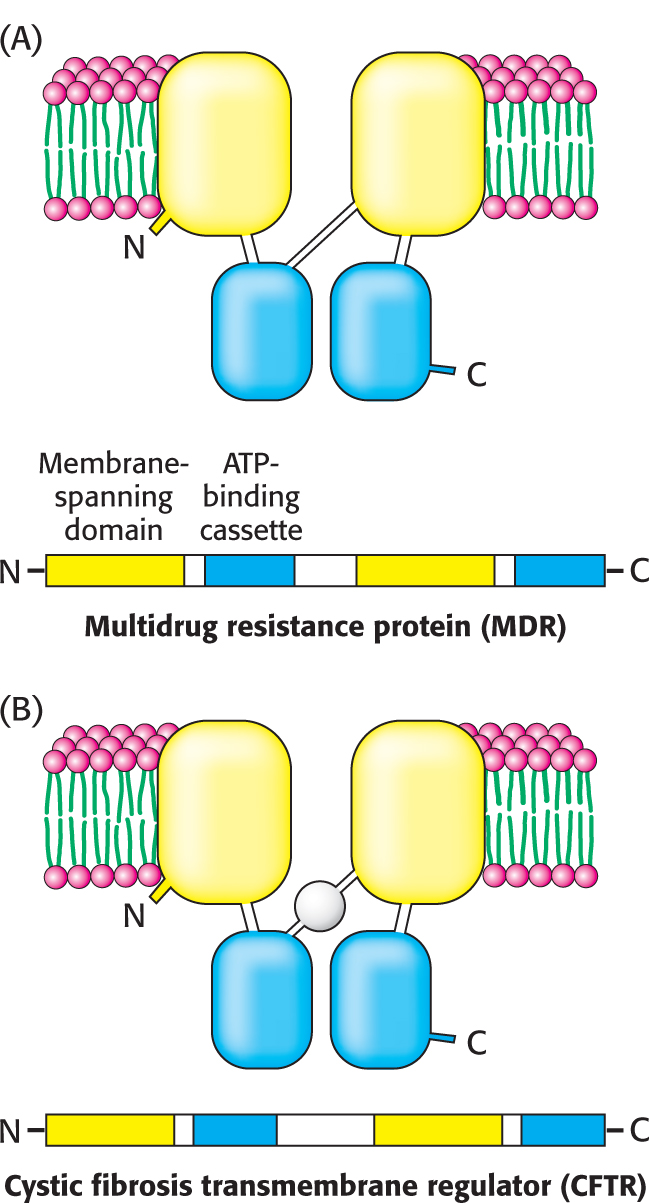
Studies of human disease revealed another large family of active-
Another example of an ABC transporter is the cystic fibrosis transmembrane conductance regulator (CFTR; Figure 12.17B). CFTR acts as an ATP-
CLINICAL INSIGHTHarlequin Ichthyosis Is a Dramatic Result of a Mutation in an ABC Transporter Protein
A number of human diseases in addition to cystic fibrosis result from defects in ABC transporter proteins. One especially startling disorder is harlequin ichthyosis, which results from a defective ABC transporter for lipids in keratinocytes, a common type of skin cell. Babies suffering from this very rare disorder are born encased in thick skin, which restricts their movement. As the skin dries out, hard diamond shaped plaques form, severely distorting facial features. The newborns usually die within a few days because of feeding difficulties, respiratory distress, or infections that are likely due to cracks in the skin.
Secondary Transporters Use One Concentration Gradient to Power the Formation of Another
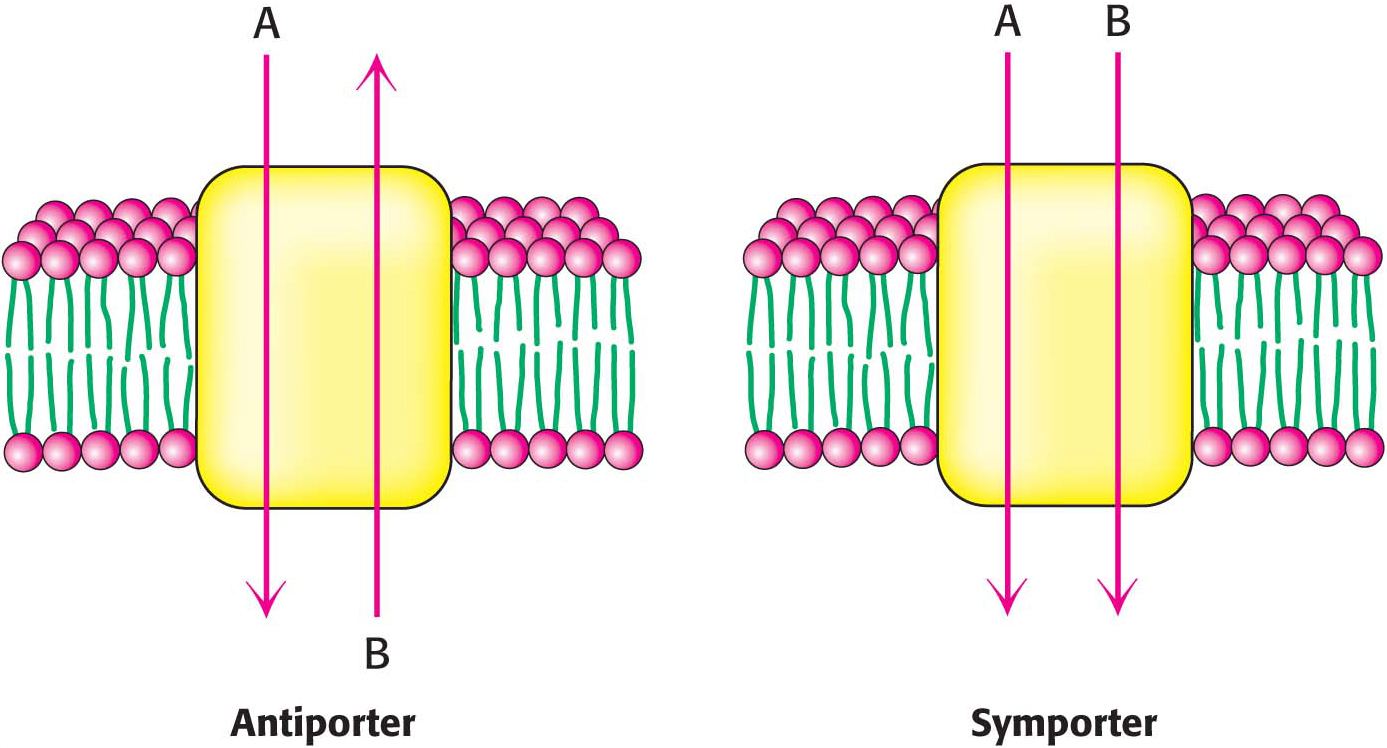
Many active-
Glucose is moved into some animal cells by the sodium-

DID YOU KNOW?
Interestingly, digitalis was used effectively long before the discovery of the Na+–K+ ATPase. In 1785, William Withering, a British physician, heard tales of an elderly woman, known as “the old woman of Shropshire,” who cured people of “dropsy” (which today would be recognized as congestive heart failure) with an extract of foxglove. Withering conducted the first scientific study of the effects of foxglove on congestive heart failure and documented its effectiveness.
CLINICAL INSIGHTDigitalis Inhibits the Na+–K+ Pump by Blocking Its Dephosphorylation
The interplay between active transport and secondary active transport is especially well illustrated by the action of the cardiotonic steroids. Heart failure can result if the muscles in the heart are not able to contract with sufficient strength to effectively pump blood. Certain steroids derived from plants, such as digitalis and ouabain, are known as cardiotonic steroids because of their ability to strengthen heart contractions. Interestingly, cardiotonic steroids exert their effect by inhibiting the Na+–K+ pump.
Digitalis is a mixture of cardiotonic steroids derived from the dried leaf of the foxglove plant Digitalis purpurea (Figure 12.20). The compound increases the force of contraction of heart muscle and is consequently a choice drug in the treatment of congestive heart failure. Inhibition of the Na+–K+ pump by digitalis means that Na+ is not pumped out of the cell, diminishing the Na+ gradient. The reduced Na+ gradient in turn affects the sodium–
Specific Channels Can Rapidly Transport Ions Across Membranes
Pumps can transport ions at rates approaching several thousand ions per second. Other membrane proteins, the passive-
Among the most important manifestations of ion-

 BIOLOGICAL INSIGHT
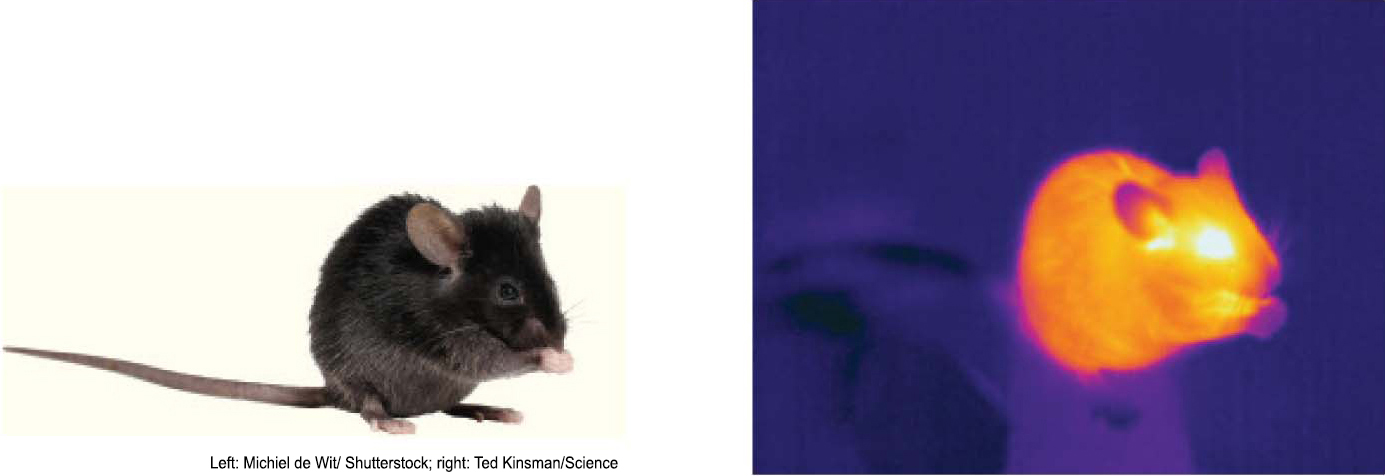
BIOLOGICAL INSIGHTVenomous Pit Vipers Use Ion Channels to Generate a Thermal Image
A large family of cation channels comprises the TRP (transient receptor potential) channels. These channels serve a host of functions in vertebrates including detecting taste, pain, and temperature. Venomous pit vipers, such as the western diamondback rattlesnake (Crotalus atrox), possess TRP channels that are activated by infrared (750 nm–
The Structure of the Potassium Ion Channel Reveals the Basis of Ion Specificity
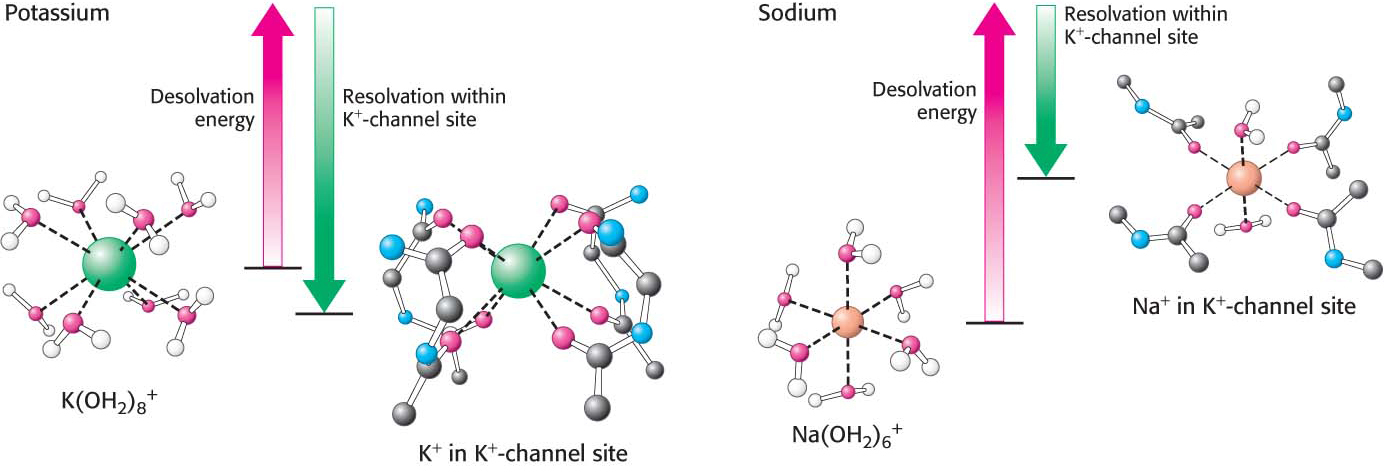
The K+ channel is one of the most extensively studied ion channels and thus provides us with a clear example of how a channel function can be both specific and rapid. Beginning from the inside of the cell, the pore starts with a diameter of approximately 10 Å and then constricts to a smaller cavity with a diameter of 8 Å (Figure 12.22). Both the opening to the outside and the central cavity of the pore are filled with water, and a K+ ion can fit in the pore without losing its shell of bound water molecules. Approximately two-

The restricted part of the pore is built from residues contributed by the two transmembrane a helices. In particular, a stretch of five amino acids within this region function as the selectivity filter that determines the preference for K+ over other ions (Figure 12.23). This region of the strand lies in an extended conformation and is oriented such that the peptide carbonyl groups are directed into the channel, in a good position to interact with the potassium ions. The potassium ion relinquishes its associated water molecules because it can bind with the oxygen atoms of the carbonyl groups of the selectivity filter.
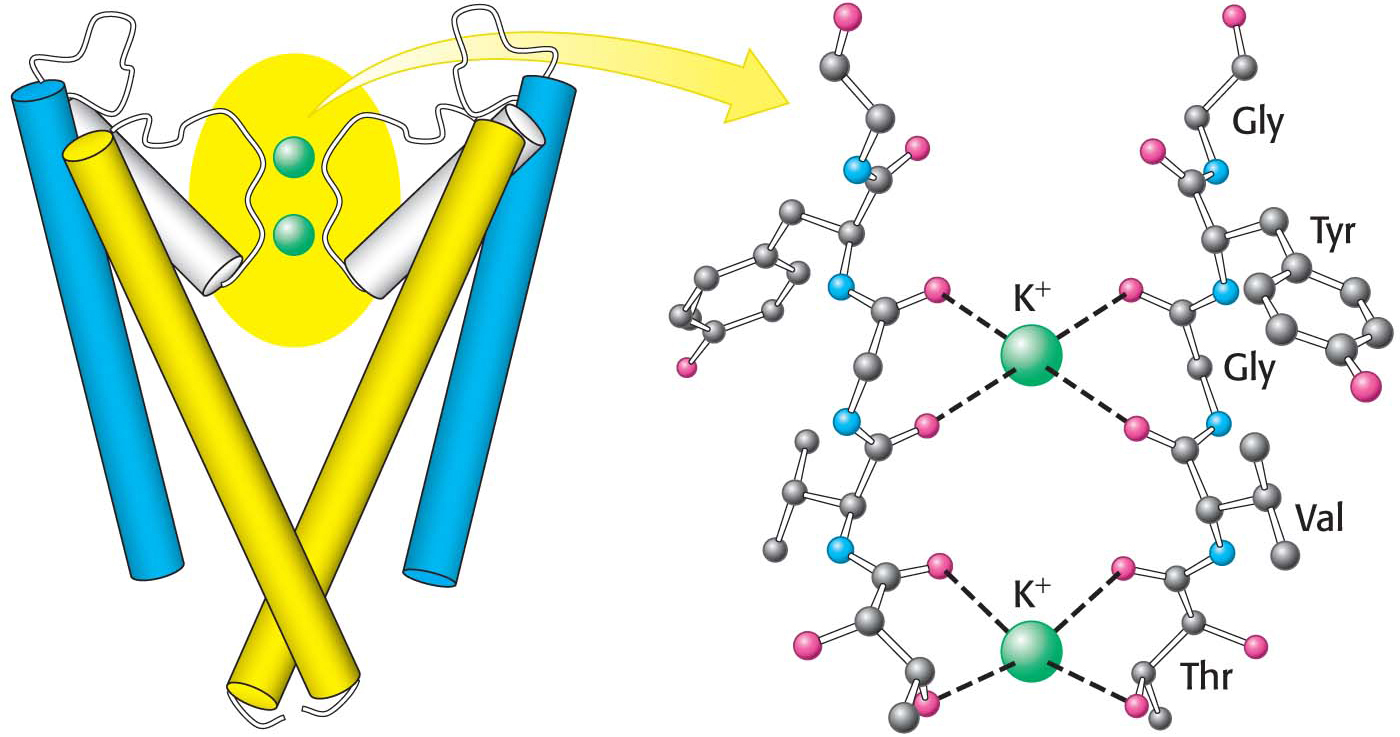
Potassium ion channels are 100-
Sodium ions are too small to react with the selectivity filter. For ions to relinquish their water molecules, other polar interactions—
The Structure of the Potassium Ion Channel Explains Its Rapid Rate of Transport
QUICK QUIZ 2
What determines the direction of flow through an ion channel?
Ion channels allow ion flow in either direction. In accordance with the Second Law of Thermodynamics, ions will flow down their concentration gradient.
The tight binding sites required for ion selectivity should slow the progress of ions through a channel, yet ion channels achieve rapid rates of ion transport. How is this apparent paradox resolved? A structural analysis of the K+ channel at high resolution provides an appealing explanation. Four K+-binding sites crucial for rapid ion flow are present in the constricted region of the K+ channel. Consider the process of ion conductance starting from inside the cell (Figure 12.25). A hydrated potassium ion proceeds into the channel and through the relatively unrestricted part of the channel. The ion then gives up its coordinated water molecules and binds to a site within the selectivity-