
17.1 Glucose Can Be Synthesized from Noncarbohydrate Precursors
✓ 3 Describe how gluconeogenesis is powered in the cell.
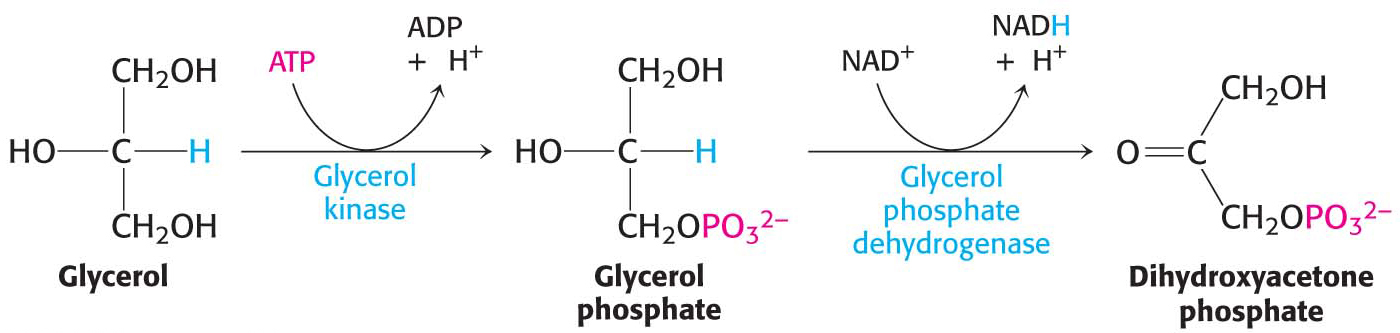
Although glucose is usually available in the environment of most organisms, this molecule is such a common fuel that a pathway exists in virtually all forms of life to synthesize it from simple precursors. The gluconeogenic pathway converts pyruvate into glucose. Noncarbohydrate precursors of glucose are first converted into pyruvate or enter the pathway at later intermediates (Figure 17.1). The major noncarbohydrate precursors are lactate, amino acids, and glycerol. Lactate is formed by active skeletal muscle through lactic acid fermentation when the rate of glycolysis exceeds the rate at which muscle can process pyruvate aerobically. Lactate is readily converted into pyruvate in the liver by the action of lactate dehydrogenase. Amino acids are derived from proteins in the diet and, during starvation, from the breakdown of proteins in skeletal muscle (Chapter 32). The hydrolysis of triacylglycerols (Chapter 27) in fat cells yields glycerol and fatty acids. Glycerol is a precursor of glucose, but animals cannot convert fatty acids into glucose for reasons that will be given later (Chapter 27). Unlike lactate and amino acids, glycerol can be metabolized by glycolysis or converted into glucose by gluconeogenesis. Glycerol may enter either the gluconeogenic or the glycolytic pathway at dihydroxyacetone phosphate.
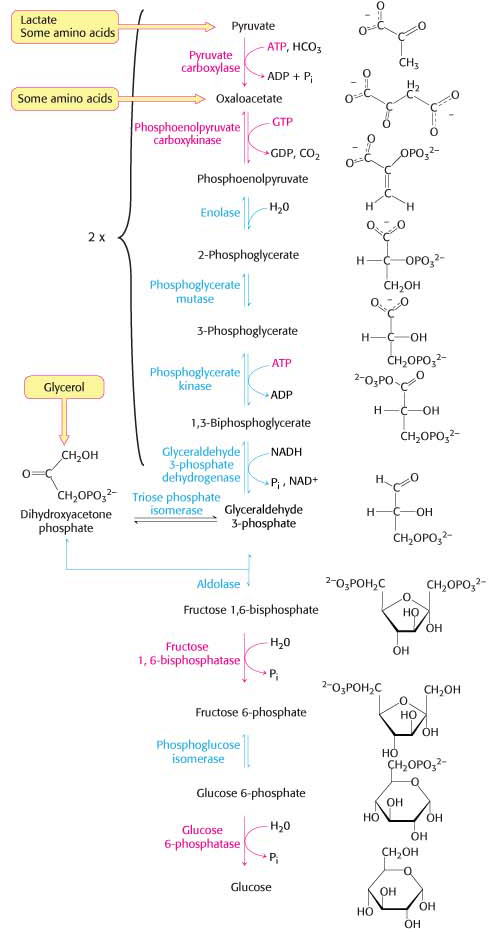
Gluconeogenesis Is Not a Complete Reversal of Glycolysis
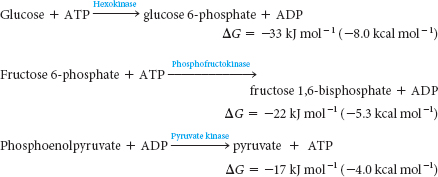
In glycolysis, glucose is converted into pyruvate; in gluconeogenesis, pyruvate is converted into glucose. However, gluconeogenesis is not a reversal of glycolysis. Several reactions must differ because the equilibrium of glycolysis lies far on the side of pyruvate formation. The actual free energy change for the formation of pyruvate from glucose is about –90 kJ mol–1 (–22 kcal mol–1) under typical cellular conditions. Most of the decrease in free energy in glycolysis takes place in the three irreversible steps catalyzed by hexokinase, phosphofructokinase, and pyruvate kinase:
In gluconeogenesis, these irreversible reactions of glycolysis must be bypassed.
The Conversion of Pyruvate into Phosphoenolpyruvate Begins with the Formation of Oxaloacetate
The first step in gluconeogenesis is the carboxylation of pyruvate to form oxaloacetate at the expense of a molecule of ATP, a reaction catalyzed by pyruvate carboxylase. This reaction occurs in the mitochondria.
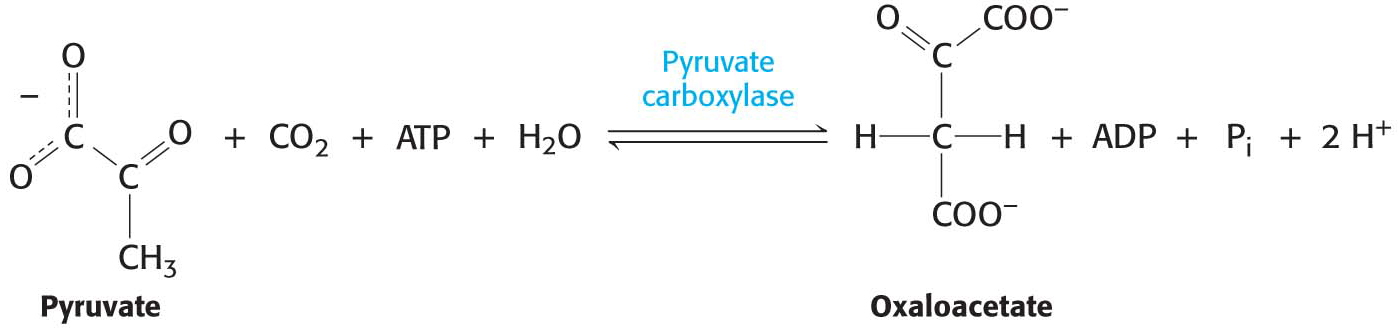
Pyruvate carboxylase requires biotin, a covalently attached prosthetic group that serves as the carrier of activated CO2. The carboxylate group of biotin is linked to the ε-amino group of a specific lysine residue by an amide bond (Figure 17.2). Recall that, in aqueous solutions, CO2 exists primarily as HCO3– with the aid of carbonic anhydrase.
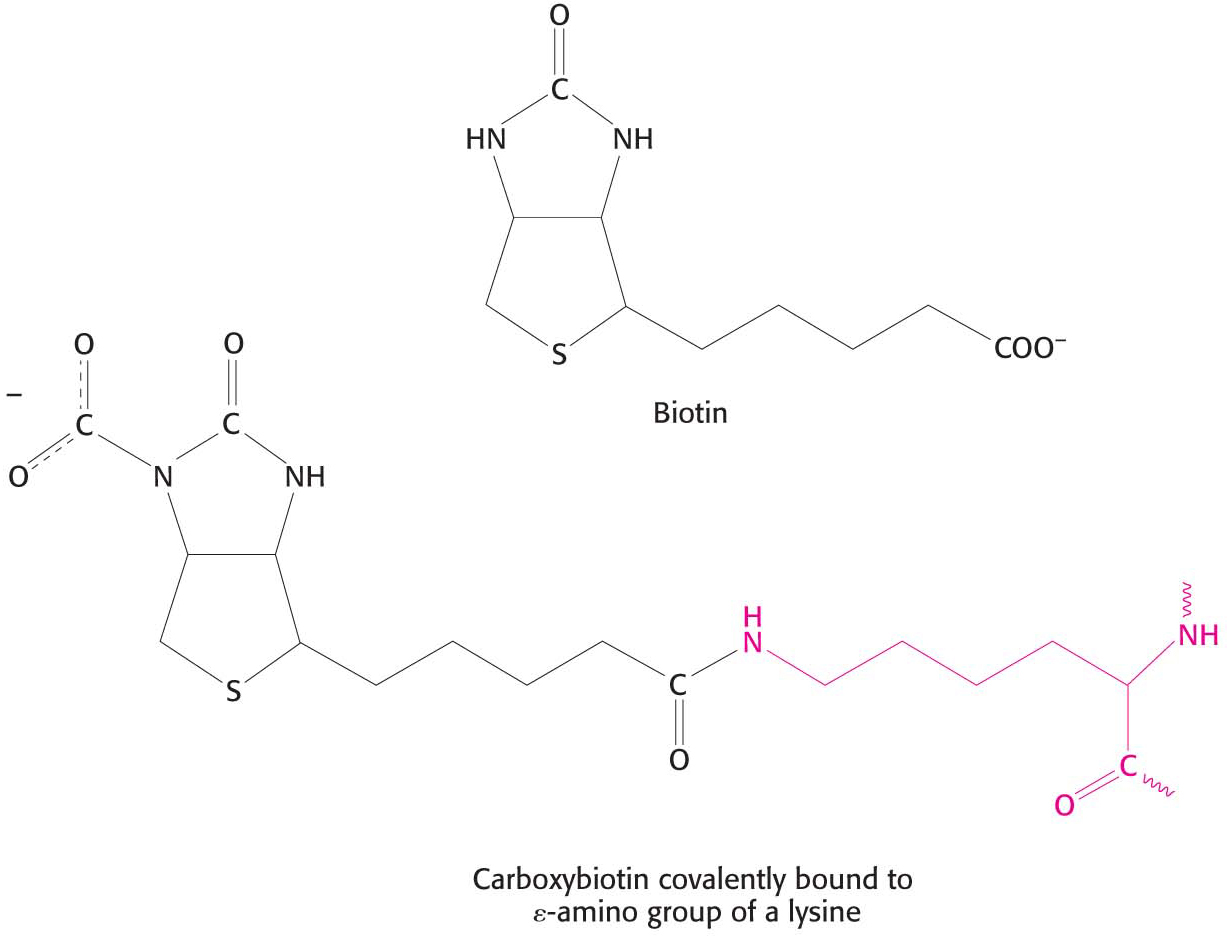
The carboxylation of pyruvate takes place in three stages:
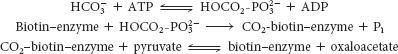
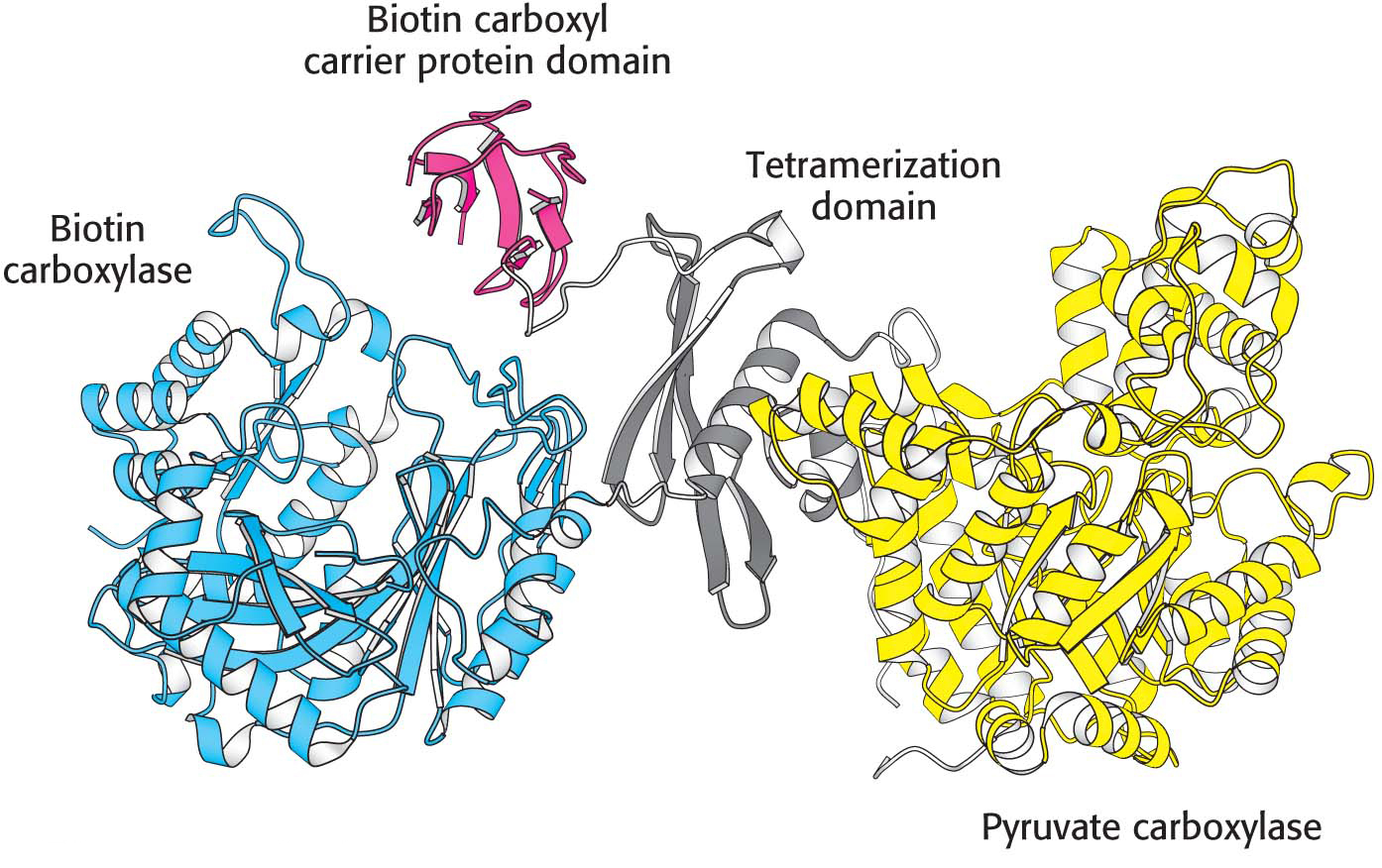
Pyruvate carboxylase functions as a tetramer composed of four identical subunits, and each subunit consists of four domains (Figure 17.3). The biotin carboxylase domain catalyzes the formation of carboxyphosphate and the subsequent attachment of CO2 to the second domain, the biotin carboxyl carrier protein (BCCP), the site of the covalently attached biotin. Once bound to CO2, BCCP leaves the biotin carboxylase active site and swings almost the entire length of the subunit (≈75Å) to the active site of the pyruvate carboxylase domain, which transfers the CO2 to pyruvate to form oxaloacetate. BCCP in one subunit interacts with the active sites on an adjacent subunit. The fourth domain facilitates the formation of the tetramer.
NUTRITION FACTS: Biotin

Also called vitamin B7, biotin is used in CO2 transfer and carboxylation reactions. Biotin deficiency is characterized by muscle pain, lethargy, anorexia, and depression. This vitamin is synthesized by microflora in the intestinal tract and can be obtained in the diet from liver, soybeans, nuts, and many other sources.
The first partial reaction of pyruvate carboxylase, the formation of carboxybiotin, depends on the presence of acetyl CoA. Biotin is not carboxylated unless acetyl CoA is bound to the enzyme. Acetyl CoA has no effect on the second partial reaction. The allosteric activation of pyruvate carboxylase by acetyl CoA is an important physiological control mechanism that will be discussed in Section 17.4.
Oxaloacetate Is Shuttled into the Cytoplasm and Converted into Phosphoenolpyruvate
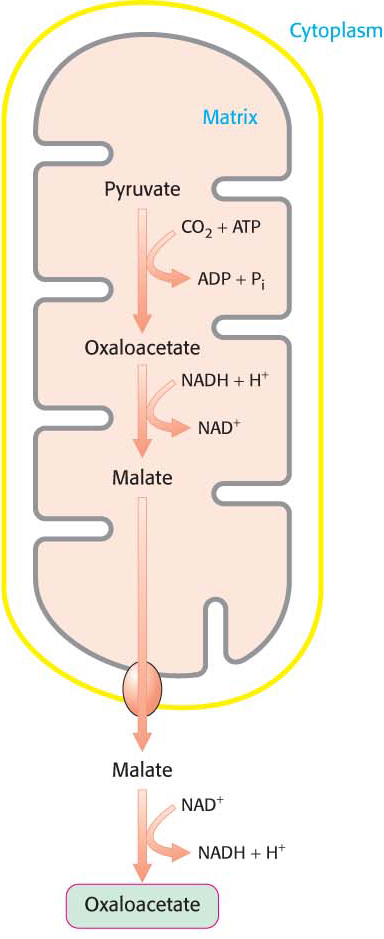
Oxaloacetate must be transported to the cytoplasm to complete the synthesis of phosphoenolpyruvate. Oxaloacetate is first reduced to malate by malate dehydrogenase. Malate is transported across the mitochondrial membrane and reoxidized to oxaloacetate by a cytoplasmic NAD+-linked malate dehydrogenase (Figure 17.4). The formation of oxaloacetate from malate also provides NADH for use in subsequent steps in gluconeogenesis. Finally, oxaloacetate is simultaneously decarboxylated and phosphorylated by phosphoenolpyruvate carboxykinase (PEPCK) to generate phosphoenolpyruvate. The phosphoryl donor is GTP. The CO2 that was added to pyruvate by pyruvate carboxylase comes off in this step:
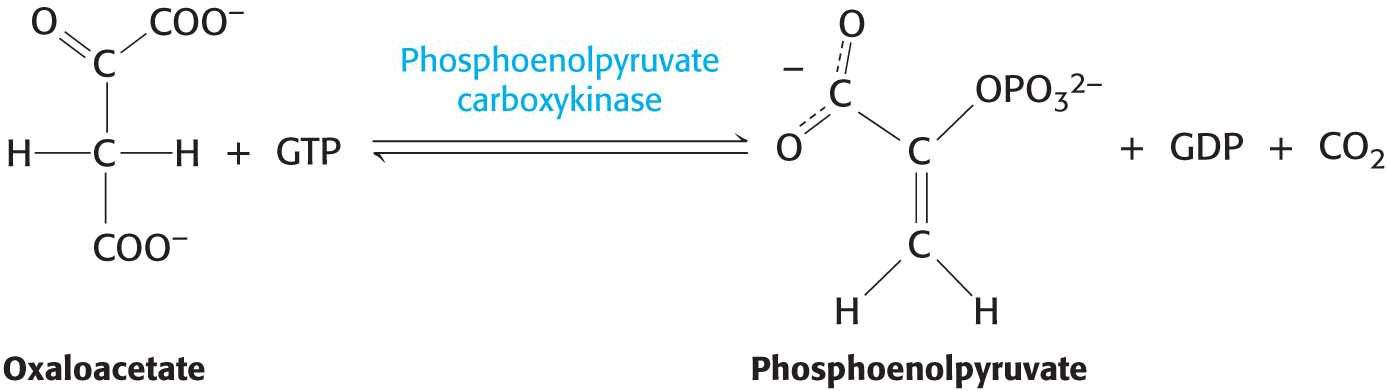
The sum of the reactions catalyzed by pyruvate carboxylase and phosphoenolpyruvate carboxykinase is
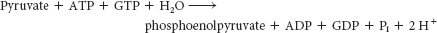
This pair of reactions bypasses the irreversible reaction catalyzed by pyruvate kinase in glycolysis.
Why is a carboxylation and a decarboxylation required to form phosphoenolpyruvate from pyruvate? Recall that, in glycolysis, the presence of a phosphoryl group traps the unstable enol isomer of pyruvate as phosphoenolpyruvate. However, the addition of a phosphoryl group to pyruvate is a highly unfavorable reaction: the ΔG°′ of the reverse of the glycolytic reaction catalyzed by pyruvate kinase is +31 kJ mol–1 (+7.5 kcal mol–1). In gluconeogenesis, the use of the carboxylation and decarboxylation steps results in a much more favorable ΔG°′. The formation of phosphoenolpyruvate from pyruvate in the gluconeogenic pathway has a ΔG°′ of +0.8 kJ mol–1 (+0.2 kcal mol–1). A molecule of ATP is used to power the addition of a molecule of CO2 to pyruvate in the carboxylation step. That CO2 is then removed to power the formation of phosphoenolpyruvate in the decarboxylation step. Decarboxylations often drive reactions that are otherwise highly endergonic. This metabolic motif is used in the citric acid cycle, the pentose phosphate pathway, and fatty acid synthesis.
The Conversion of Fructose 1,6-bisphosphate into Fructose 6-phosphate and Orthophosphate Is an Irreversible Step
The newly formed phosphoenolpyruvate is then metabolized by the enzymes of glycolysis but in the reverse direction. These reactions are near equilibrium under intracellular conditions; so, when conditions favor gluconeogenesis, the reverse reactions will take place until the next irreversible step is reached. This step is the hydrolysis of fructose 1,6-

Fructose 1,6-
The Generation of Free Glucose Is an Important Control Point
The fructose 6-
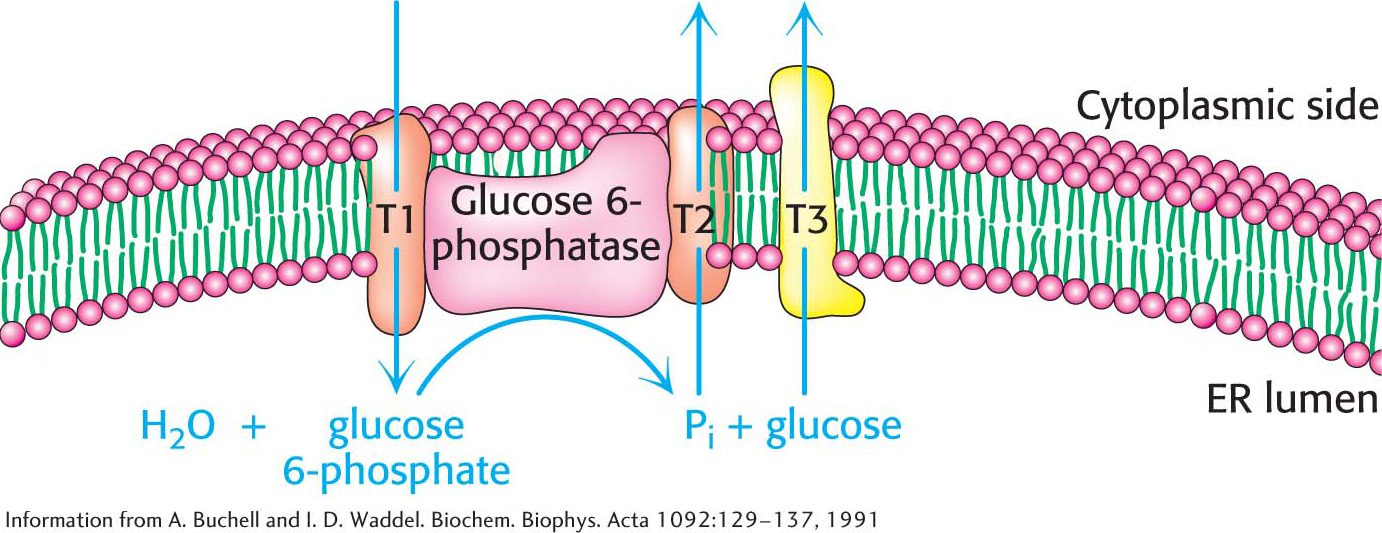
Six High-Transfer-Potential Phosphoryl Groups Are Spent in Synthesizing Glucose from Pyruvate
As we have seen in the preceding sets of reactions, the formation of glucose from pyruvate is energetically unfavorable unless it is coupled to reactions that are favorable. Compare the stoichiometry of gluconeogenesis with that of the reverse of glycolysis. The stoichiometry of gluconeogenesis is
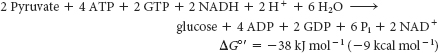
In contrast, the stoichiometry for the reversal of glycolysis is
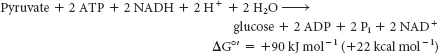
QUICK QUIZ 1
What barrier prevents glycolysis from simply running in reverse to synthesize glucose? How is this barrier overcome in gluconeogenesis?
The reverse of glycolysis is highly endergonic under cellular conditions. The expenditure of six NTP molecules in gluconeogenesis renders gluconeogenesis exergonic.
Note that six nucleoside triphosphate molecules are hydrolyzed to synthesize glucose from pyruvate in gluconeogenesis, whereas only two molecules of ATP are generated in glycolysis in the conversion of glucose into pyruvate. Thus, the extra cost of gluconeogenesis is four high-