
1.4 Membranes Define the Cell and Carry Out Cellular Functions
✓ 3 Identify the key features that differentiate eukaryotic cells from prokaryotic cells.
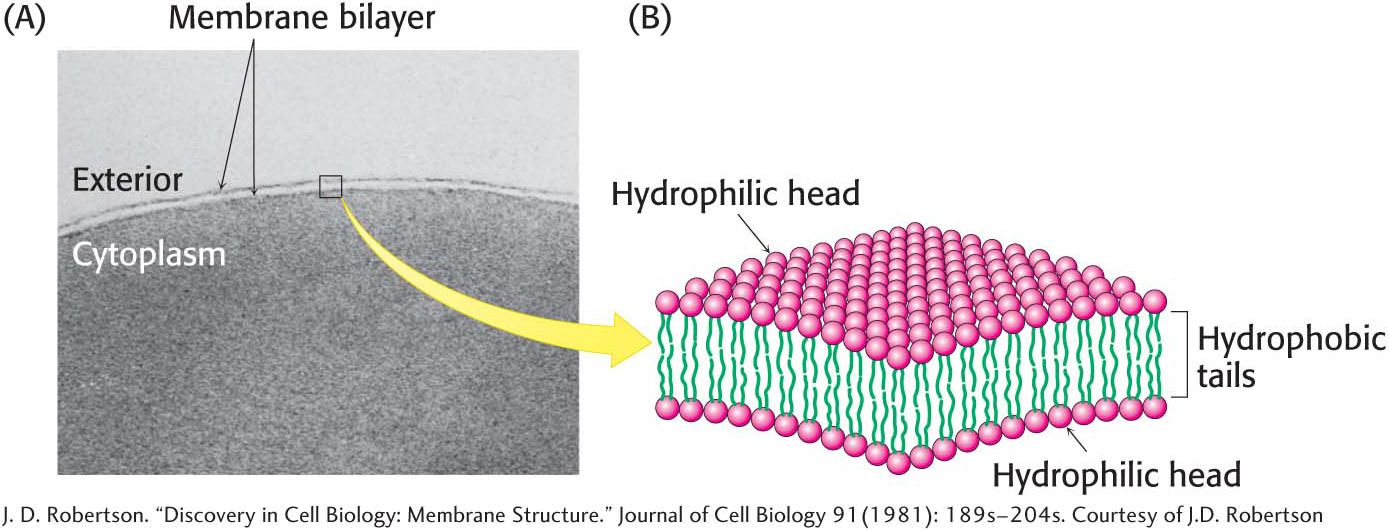
The cell is the basic unit of life. Cells grow, replicate, and interact with their environment. Living organisms can be as simple as a single cell or as complex as a human body, which is composed of approximately 100 trillion cells. Every cell is delineated by a membrane that separates the inside of the cell from its environment. A membrane is a lipid bilayer: two layers of lipids organized with their hydrophobic chains interacting with one another and the hydrophilic head groups interacting with the environment (Figure 1.9).
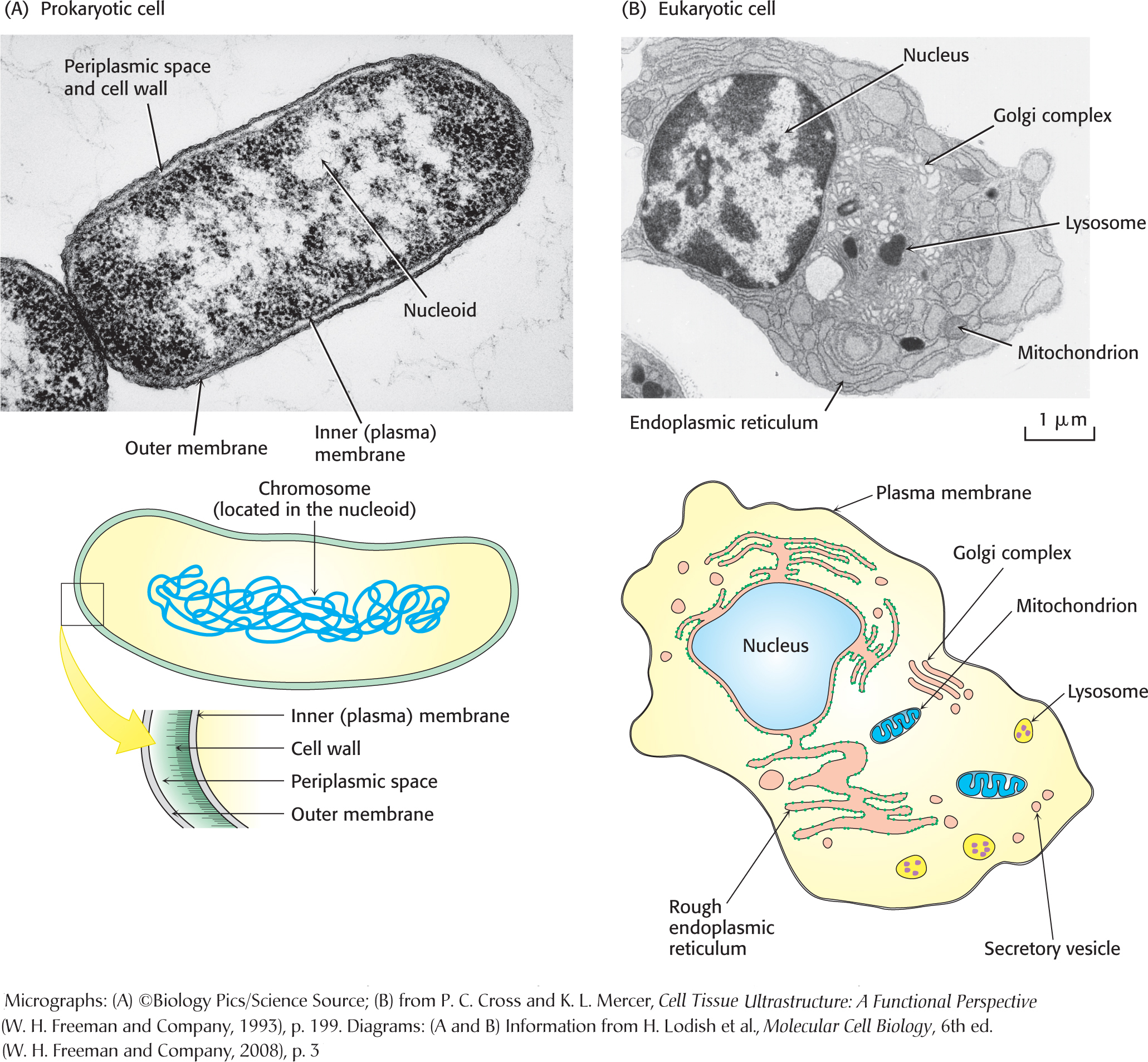
There are two basic types of cells: eukaryotic cells and prokaryotic cells (Figure 1.10). The main difference between the two is the existence of membrane-
Regardless of cell type—
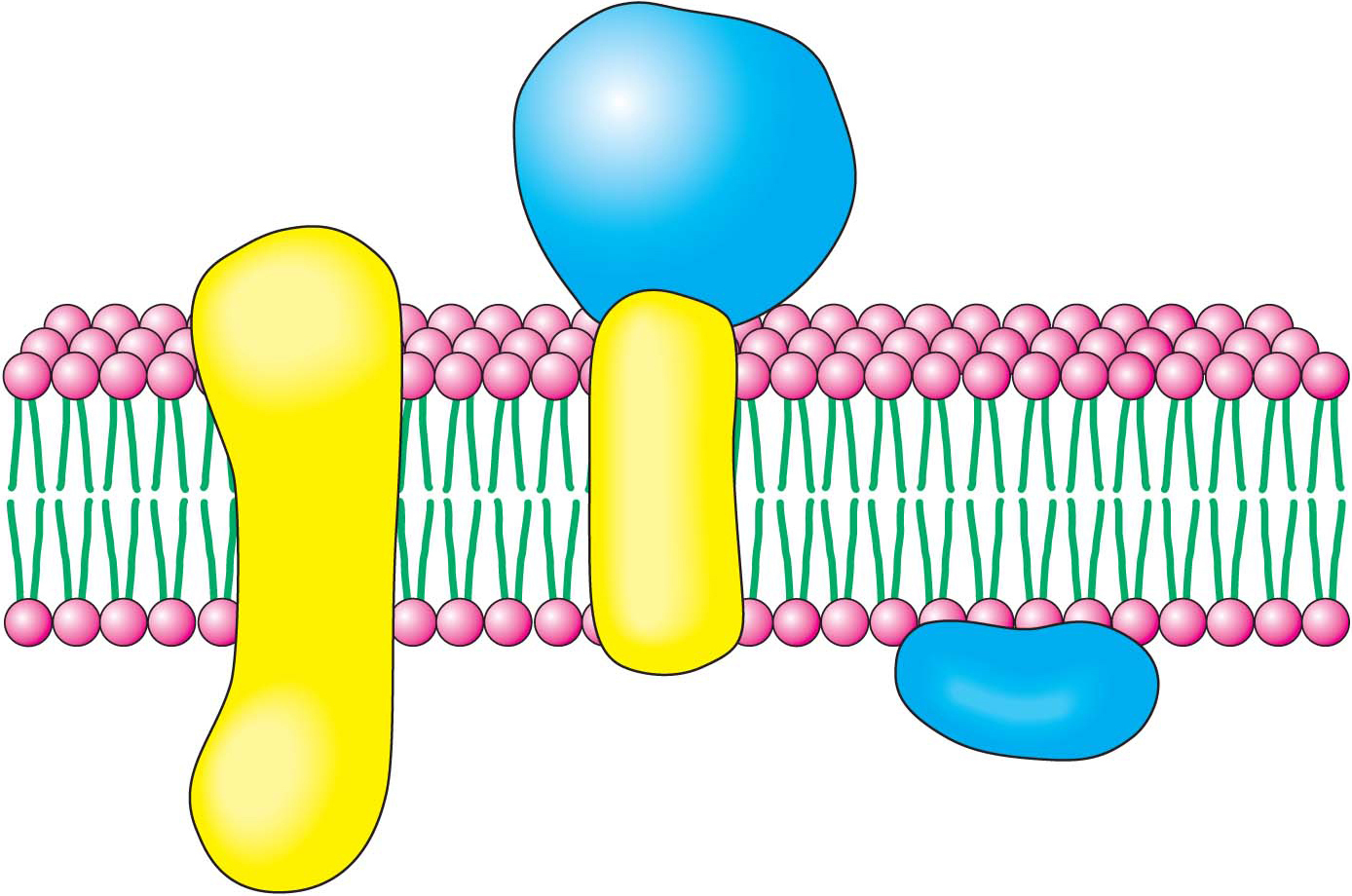
The plasma membraneThe plasma membrane separates the inside of the cell from the outside, one cell from another cell. This membrane is impermeable to most substances, even to substances such as fuels, building blocks, and signal molecules that must enter the cell. Consequently, the barrier function of the membrane must be mitigated to permit the entry and exit of molecules and information. In other words, the membrane must be rendered semipermeable but in a very selective way. This selective permeability is the work of proteins that are embedded in the plasma membrane or associated with it (Figure 1.11). These proteins facilitate the entrance of fuels, such as glucose, and building blocks, such as amino acids, and they transduce information—
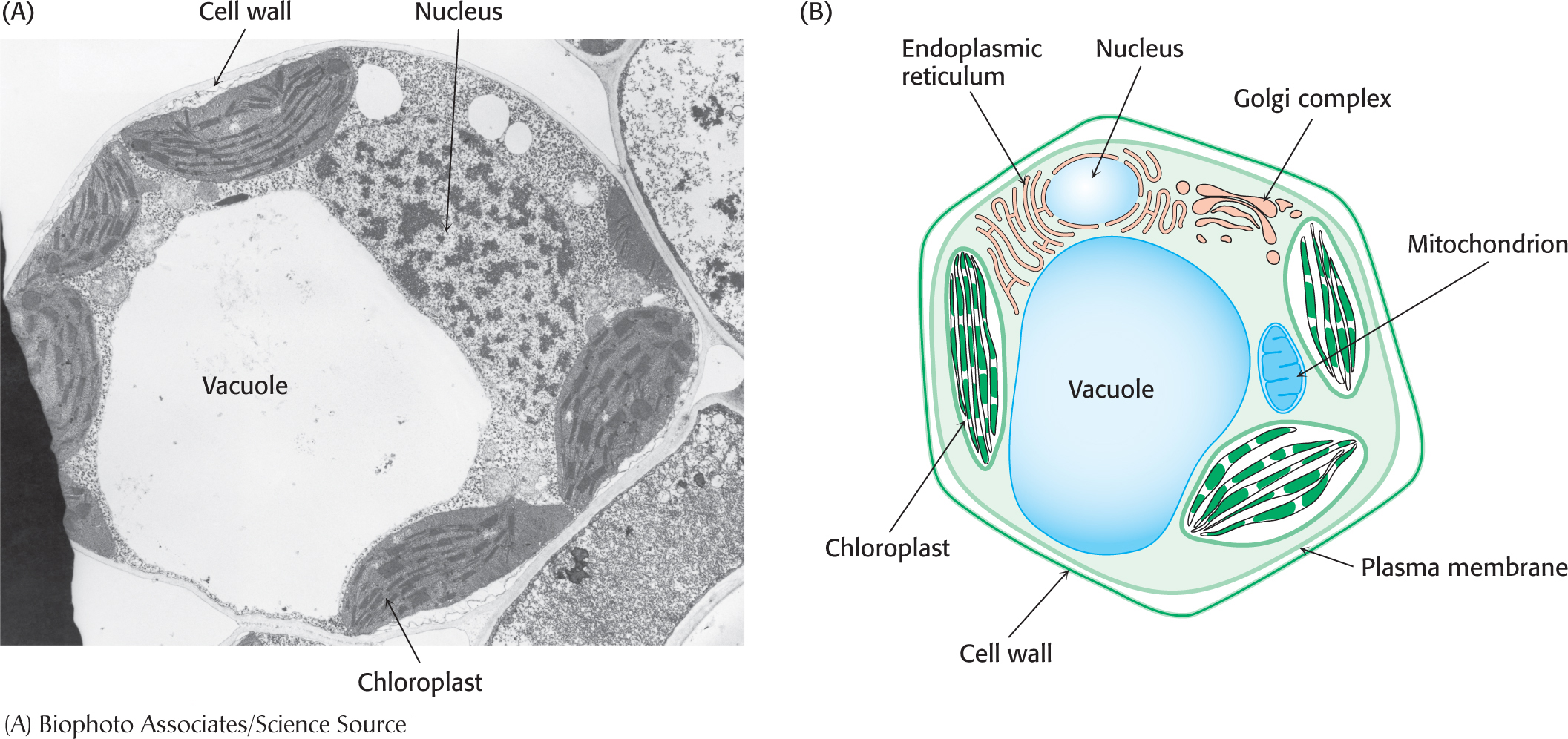
The plant cell wallThe plasma membrane of a plant is itself surrounded by a cell wall (Figure 1.12). The cell wall is constructed largely from cellulose, a long, linear polymer of glucose molecules. Cellulose molecules interact with one another as well as with other cell-
The cytoplasmThe inner substance of the cell, the material that is surrounded by the plasma membrane, is called the cytoplasm. The cytoplasm is the site of a host of biochemical processes, including the initial stage of glucose metabolism, fatty acid synthesis, and protein synthesis. Formerly, the cytoplasm was believed to be a “soup” of important biomolecules, but it is becoming increasingly clear that the biochemistry of the cytoplasm is highly organized by a network of structural filaments called the cytoskeleton. In many eukaryotes, the cytoskeleton is a network of three kinds of protein fibers—
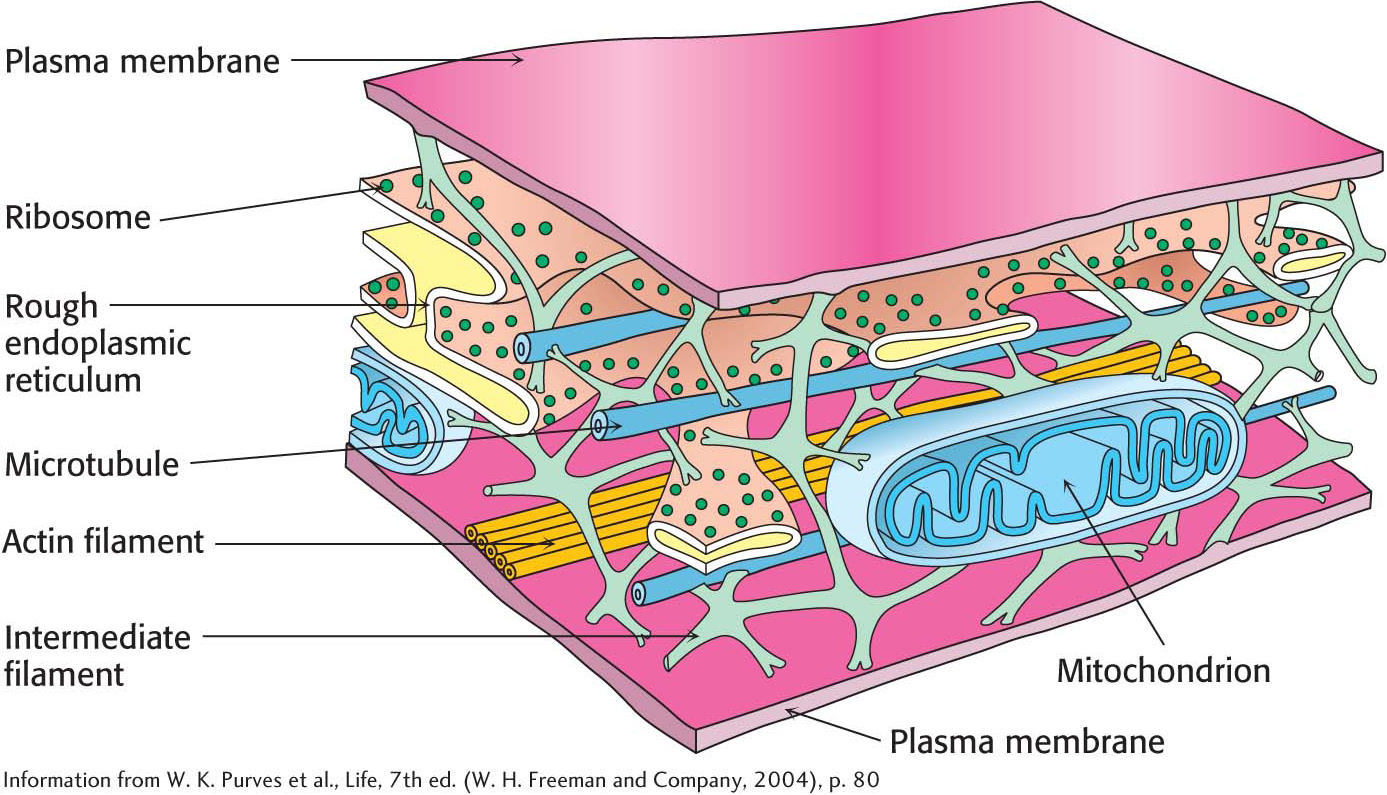
Biochemical Functions Are Sequestered in Cellular Compartments
A key difference between eukaryotic cells and prokaryotic cells is the presence of a complex array of intracellular, membrane-
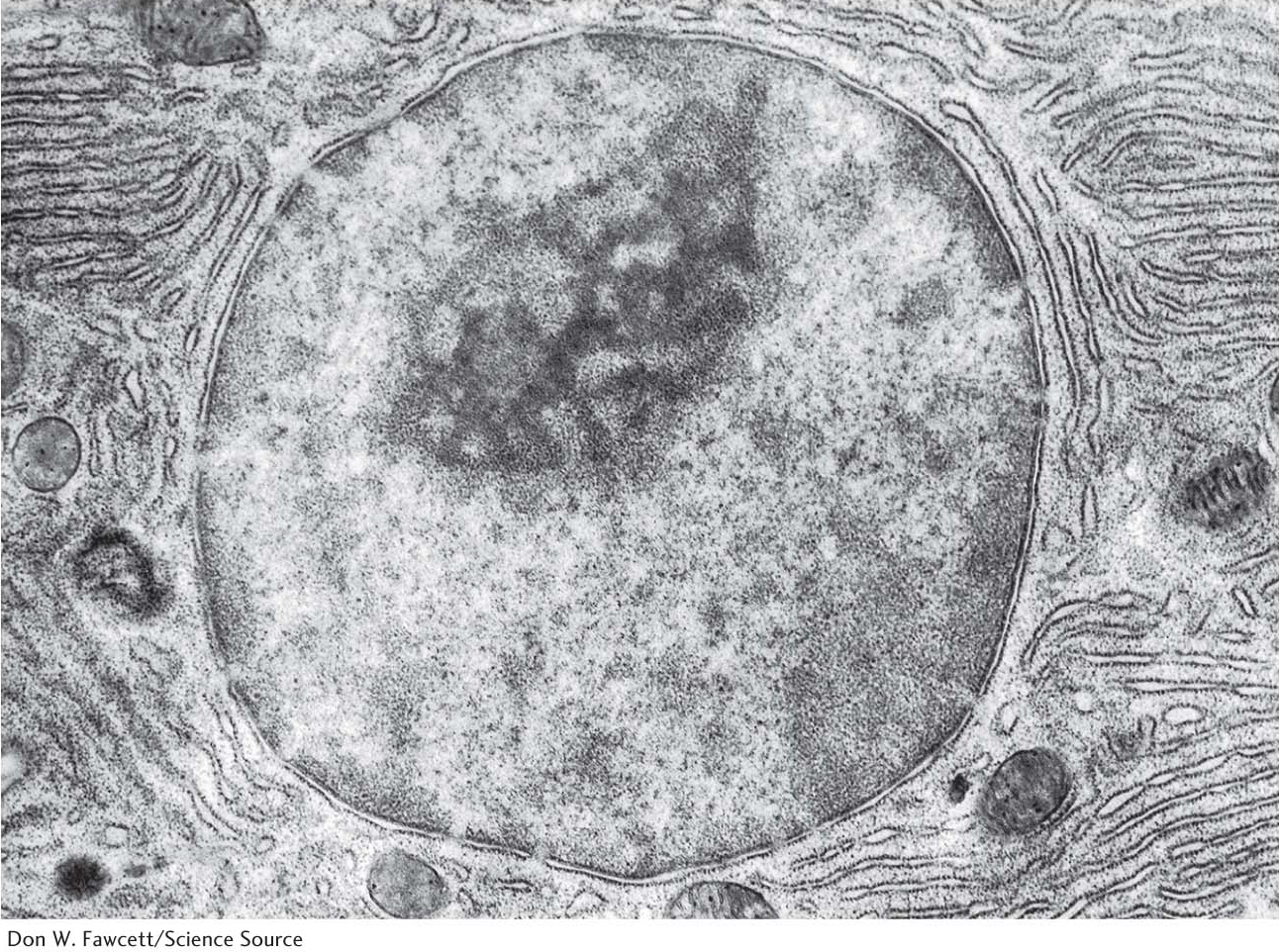
The nucleusThe largest organelle is the nucleus, which is a double-
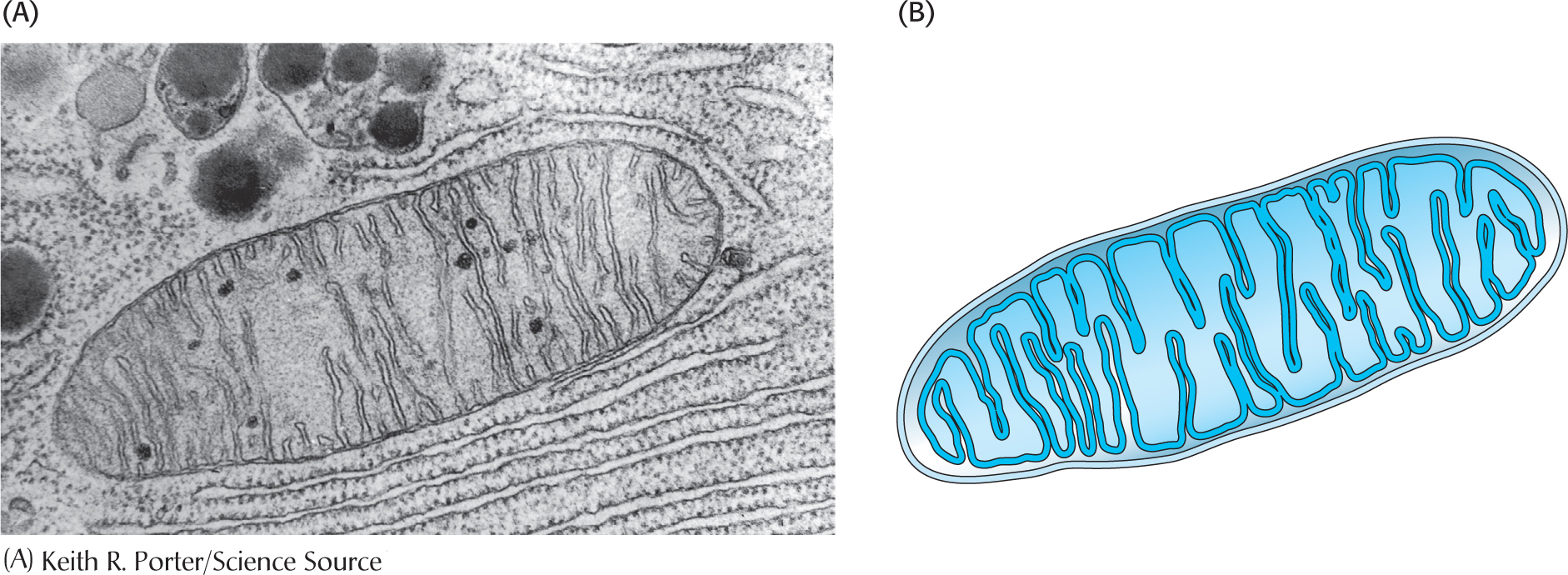
The mitochondrionThe mitochondrion (plural, mitochondria) has two membranes—
The chloroplastAnother double-
Some Organelles Process and Sort Proteins and Exchange Material with the Environment
Let us briefly examine other eukaryotic organelles (Figure 1.10B) in the context of how they cooperate with one another to perform vital biochemical tasks.
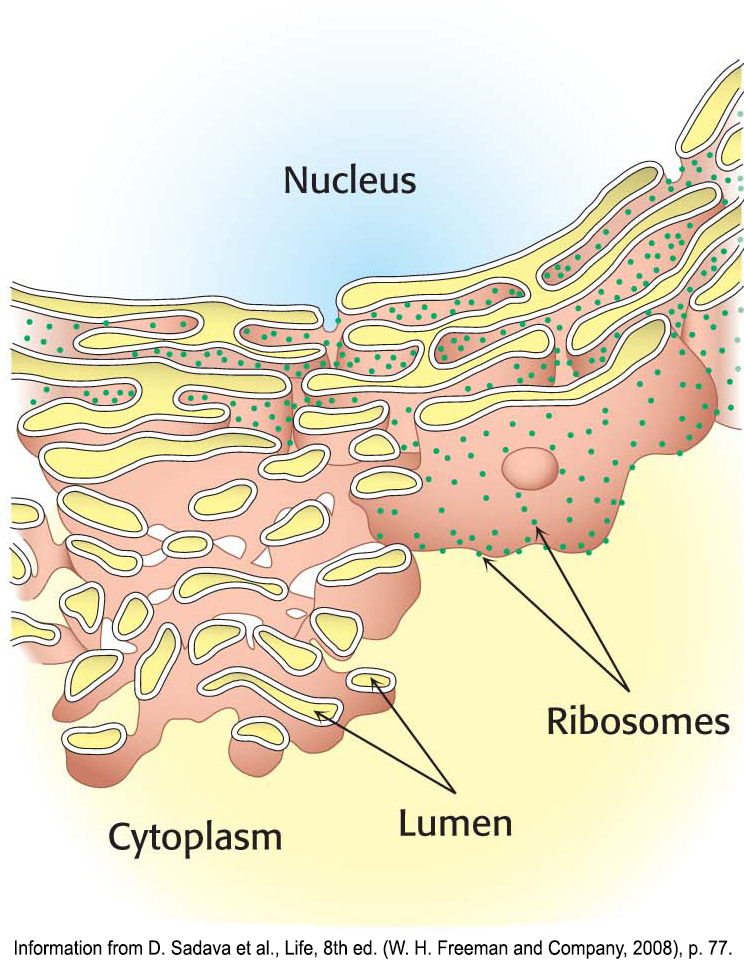
The endoplasmic reticulumThe endoplasmic reticulum is a series of membranous sacs. Many biochemical reactions take place on the cytoplasmic surface of these sacs as well as in their interiors, or lumens. The endoplasmic reticulum comes in two types: the smooth endoplasmic reticulum (smooth ER, or SER) and the rough endoplasmic reticulum (rough ER, or RER), as illustrated in Figure 1.16 (also Figure 1.10). The smooth endoplasmic reticulum plays a variety of roles, but an especially notable role is the processing of exogenous chemicals (chemicals originating outside the cell) such as drugs. The more drugs, including alcohol, ingested by an organism, the greater the quantity of smooth endoplasmic reticulum in the liver.
The rough endoplasmic reticulum appears rough because ribosomes are attached to the cytoplasmic side (Figure 1.14). Ribosomes that are free in the cytoplasm take part in the synthesis of proteins for use inside the cell. Ribosomes attached to the rough endoplasmic reticulum synthesize proteins that will either be inserted into cellular membranes or be secreted from the cell.
Proteins synthesized on the rough endoplasmic reticulum are transported into the lumen of the endoplasmic reticulum during the process of translation. Inside the lumen of the rough endoplasmic reticulum, a protein folds into its final three-
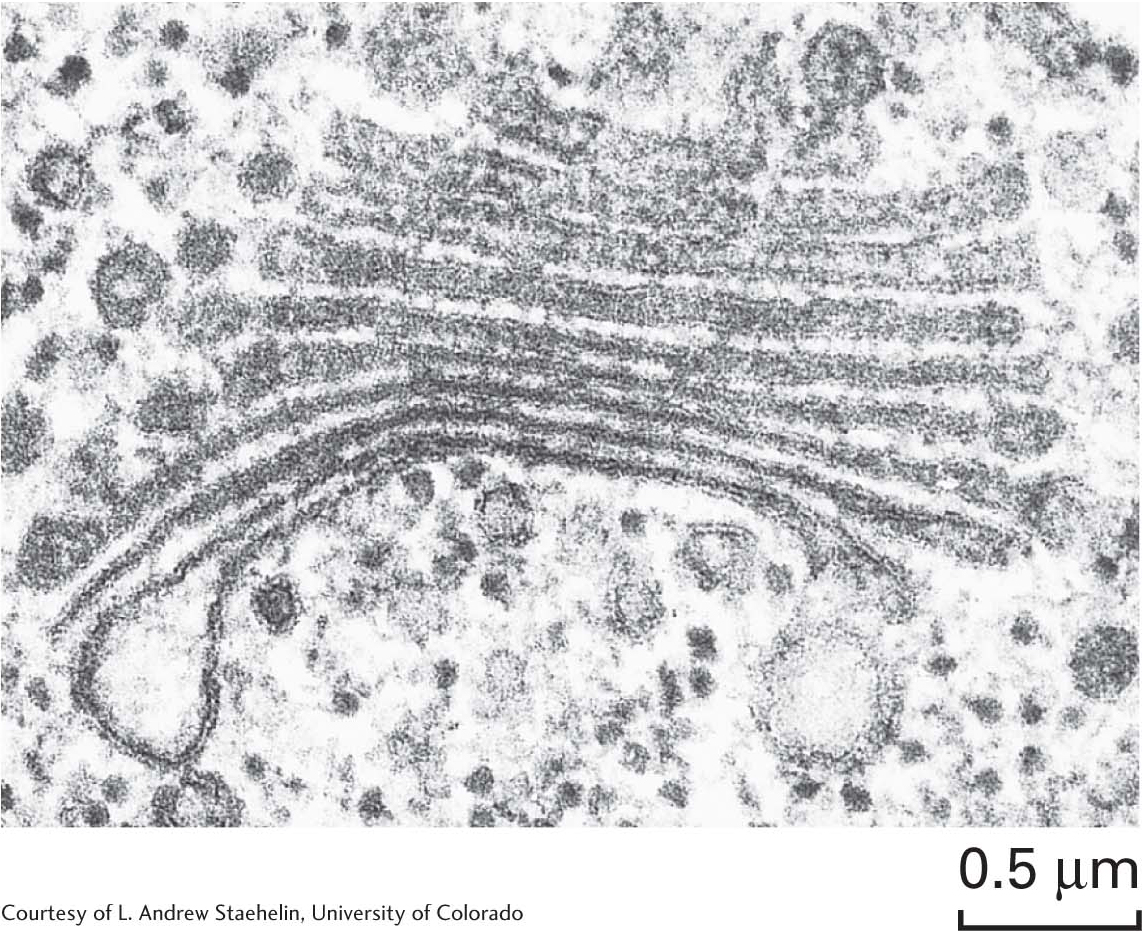
The Golgi complexThe transport vesicles from the rough endoplasmic reticulum are carried to the Golgi complex—a series of stacked membranes—
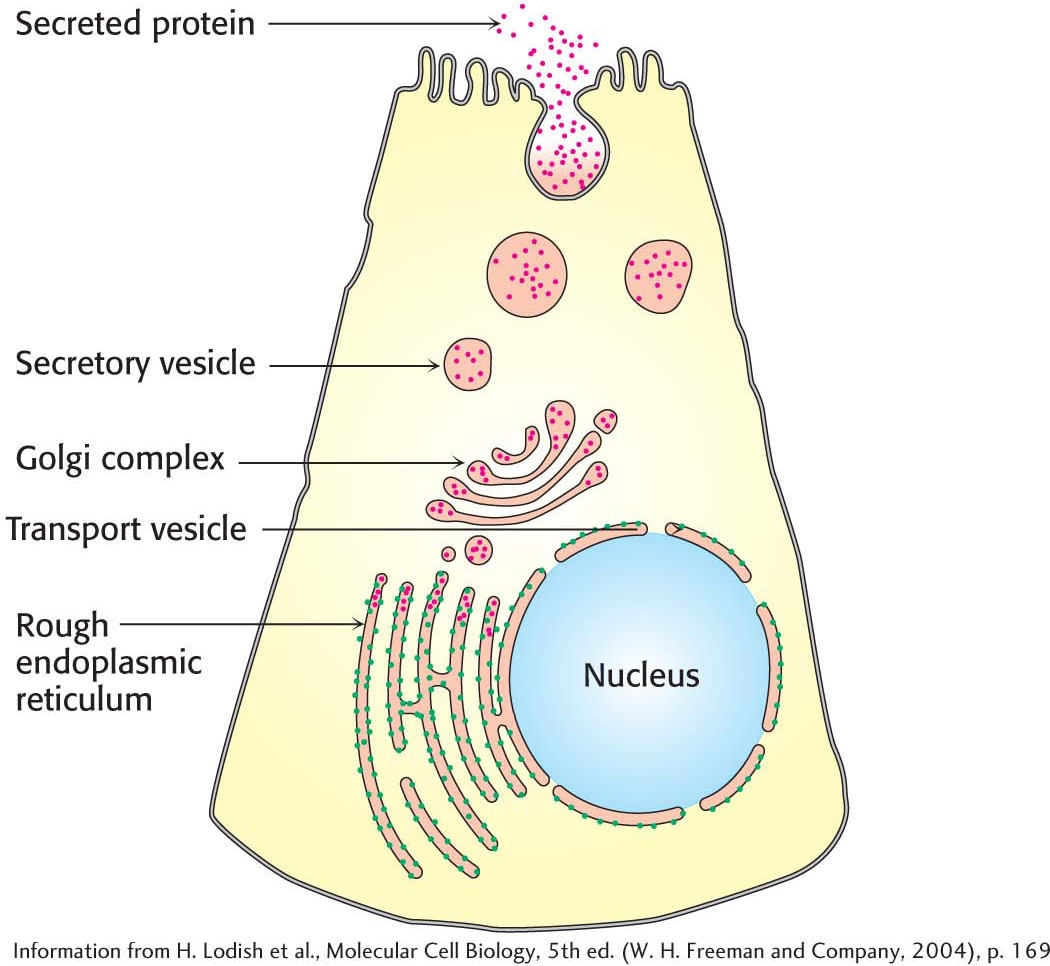
Secretory granulesA secretory granule, or zymogen granule, is formed when a vesicle filled with the proteins destined for secretion buds off the Golgi complex. The granule is directed toward the cell membrane. When the proper signal is received, the secretory granule fuses with the plasma membrane and dumps its cargo into the extracellular environment, a process called exocytosis (Figure 1.18).
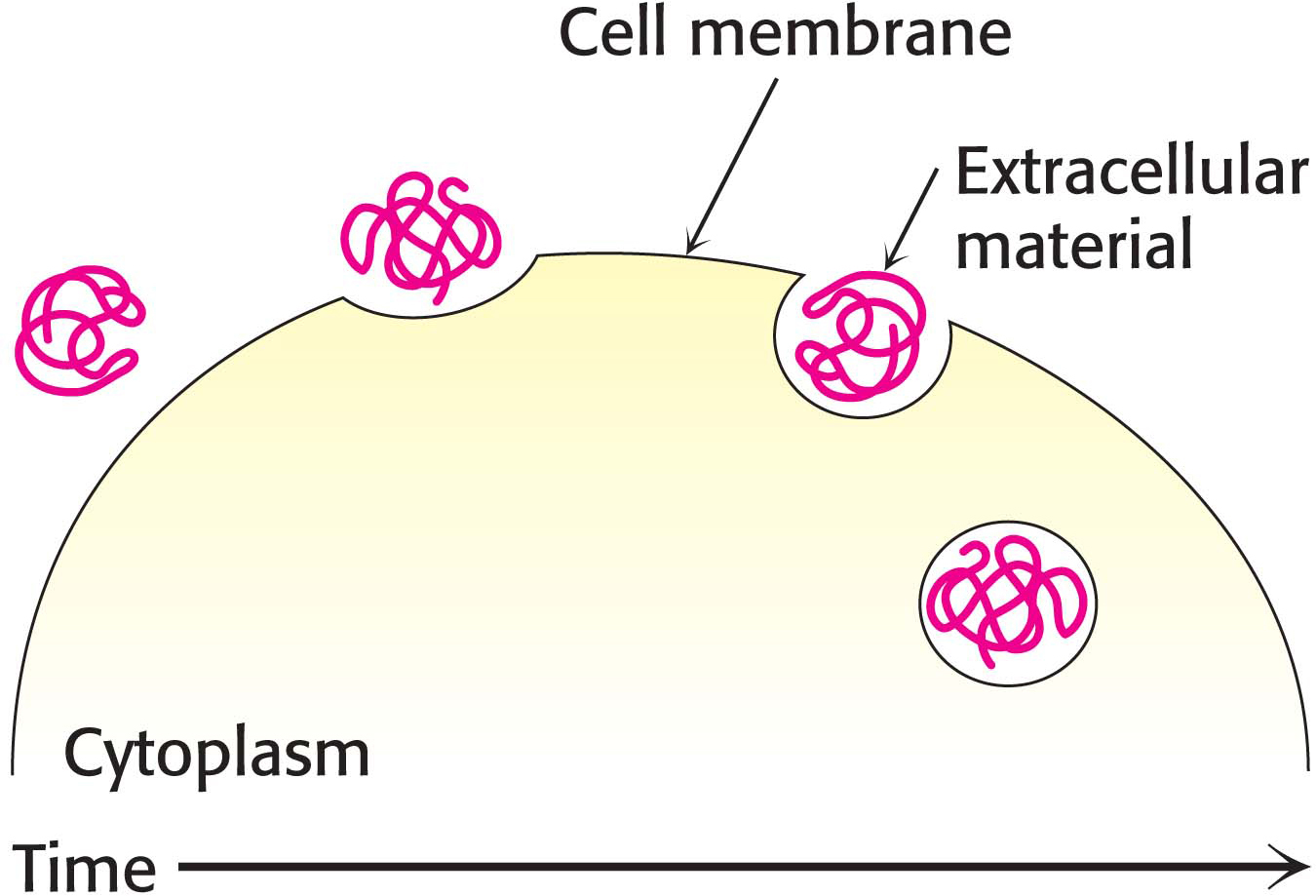
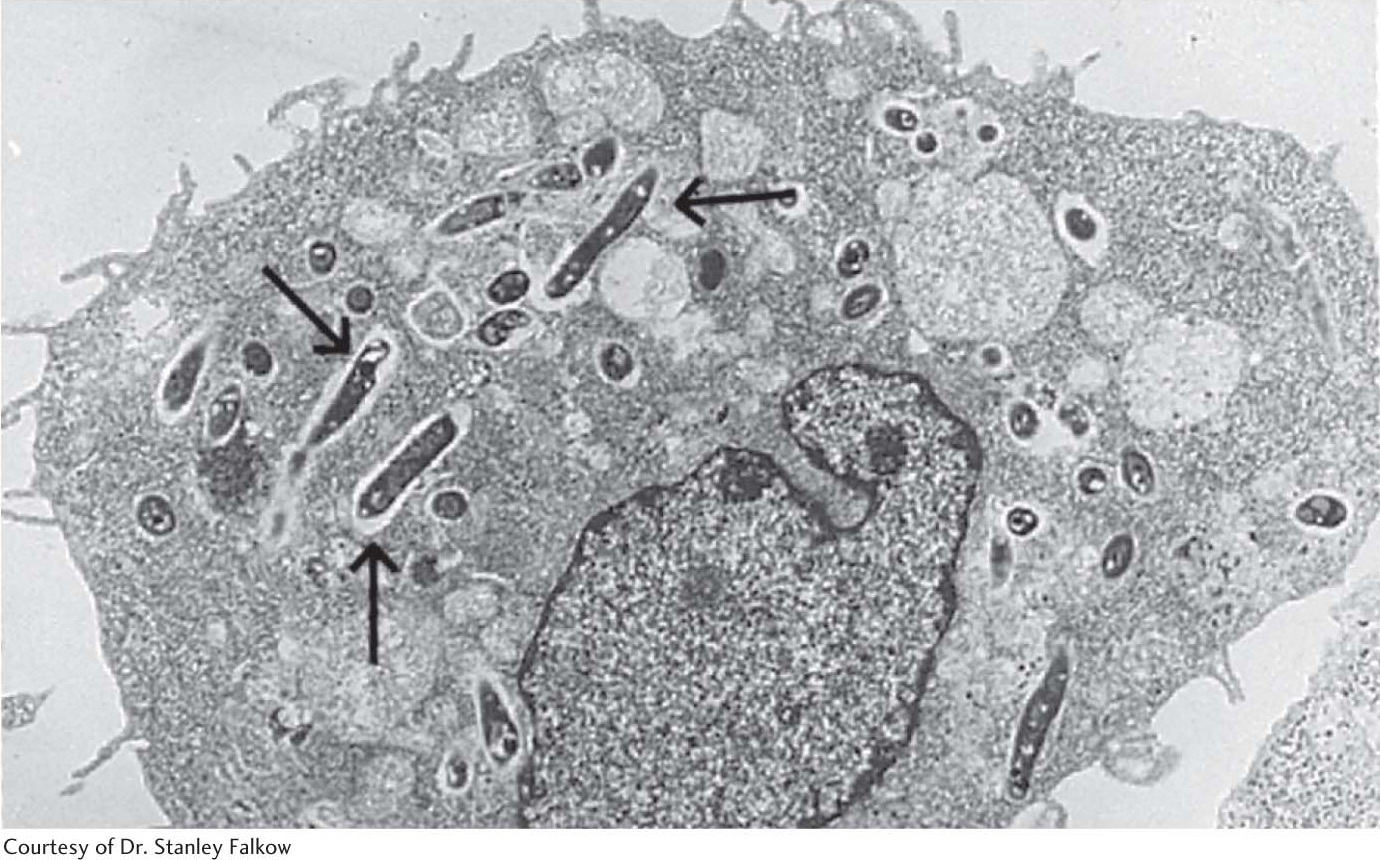
The endosomeMaterial is taken into the cell when the plasma membrane invaginates and buds off to form an endosome (not shown in Figure 1.10B). This process is called endocytosis, which is the opposite of exocytosis. Endocytosis is used to bring important biochemicals such as iron ions, vitamin B12, and cholesterol into the cell. Endocytosis takes place through small regions of the membrane, such as when a protein is taken into the cell (Figure 1.19). Alternatively, large amounts of material can also be taken into the cell. When large amounts of material are taken into the cell, the process is called phagocytosis. Figure 1.20 shows an immune-
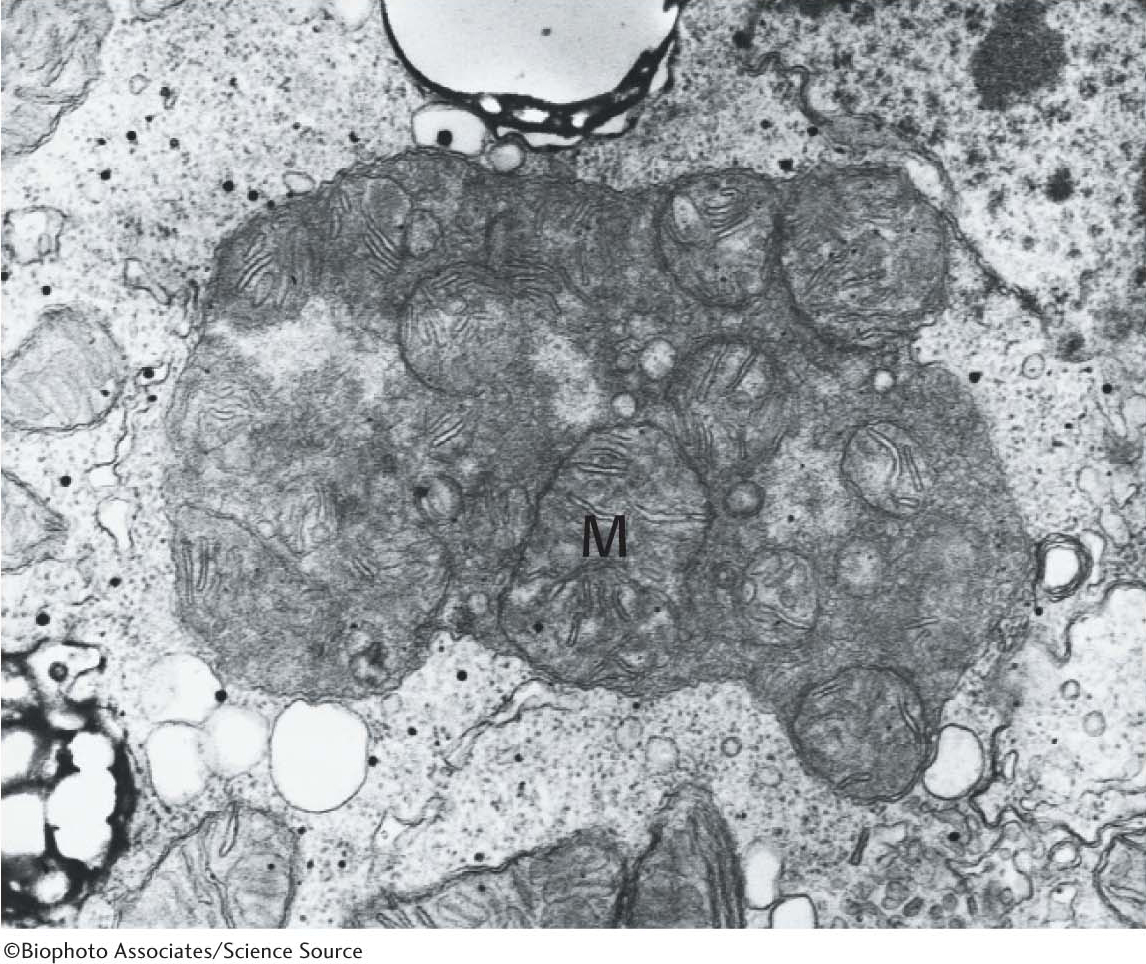
LysosomesThe lysosome is an organelle that contains a wide array of digestive enzymes. Lysosomes form in a manner analogous to the formation of secretory granules, but lysosomes fuse with endosomes instead of the cell membrane. After fusion has taken place, the lysosomal enzymes digest the material, releasing small molecules that can be used as building blocks or fuel by the cell. Lysosomes do not just degrade extracellular material, however. Another role is the digestion of damaged intracellular organelles (Figure 1.21).
QUICK QUIZ 2
Name three organelles or structures found in plant cells but not in animal cells.
Chloroplasts, vacuoles, and cell wall.
Plant vacuolesAnother organelle unique to plant cells, in addition to the chloroplast, is a large vacuole. In some plant cells, this single-
 CLINICAL INSIGHT
CLINICAL INSIGHTDefects in Organelle Function May Lead to Disease
Many pathological conditions arise due to malfunctions in various organelles. For instance, familial hypercholesterolemia, a disease in which children as young as 6 years old die of heart attacks, is caused by the inefficient endocytosis of cholesterol from the blood. The resulting high levels of cholesterol in the blood result in heart attacks. Tay–
Cellular organization attests to the high information content of the cell. But this brief overview has only touched the surface of the information processing that must take place to construct something as sophisticated as a cell. In the rest of this textbook, we will examine the biochemical energy and information pathways that construct and maintain living systems.