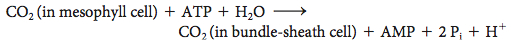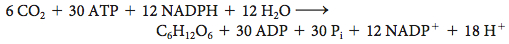23.2The Calvin Cycle Is Regulated by the Environment
✓ 5 Describe how the light reactions and the Calvin cycle are coordinated.
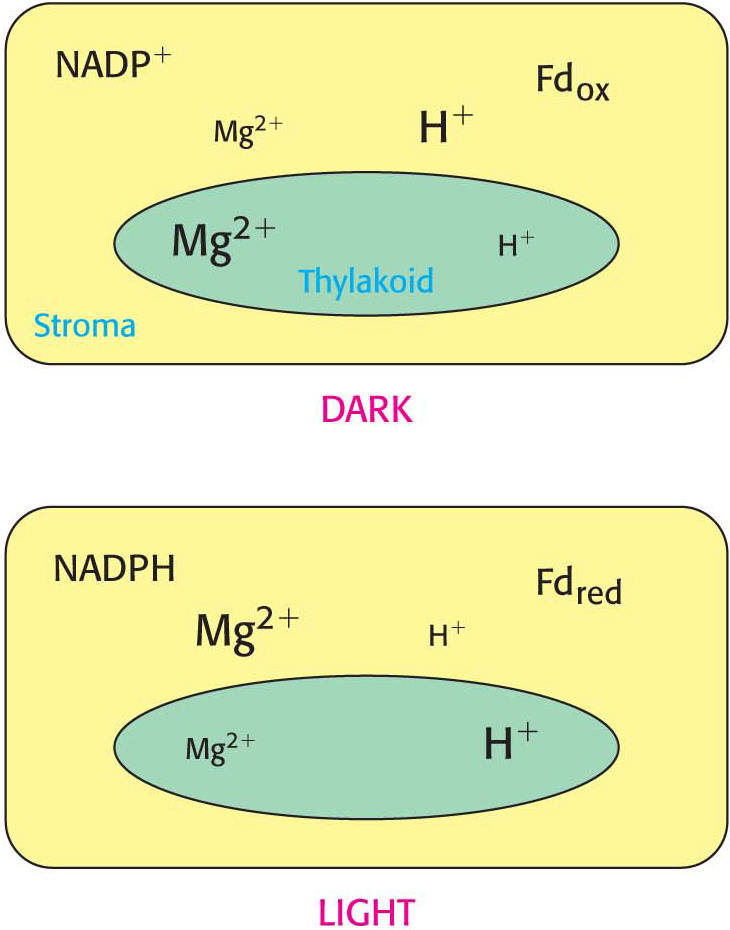
How do the light reactions communicate with the dark reactions to regulate the crucial process of fixing CO2 into biomolecules? The principal means of regulation is alteration of the stromal environment by the light reactions. The light reactions lead to an increase in pH and stromal concentrations of Mg2+, NADPH, and reduced ferredoxin—all of which contribute to the activation of certain Calvin-cycle enzymes (Figure 23.11).
Page 435
Figure 23.11 Light regulation of the Calvin cycle. The light reactions of photosynthesis transfer electrons out of the thylakoid lumen into the stroma, and they transfer H+ from the stroma into the thylakoid lumen. As a consequence of these processes, the concentrations of NADPH, reduced ferredoxin (Fd), and Mg2+ in the stroma are higher in the light than in the dark, whereas the concentration of H+ is lower in the light. Each of these concentration changes helps couple the Calvin-cycle reactions to the light reactions.
As stated earlier, the rate-limiting step in the Calvin cycle is the carboxylation of ribulose 1,5-bisphosphate to form two molecules of 3-phosphoglycerate. This step is catalyzed by rubisco, a very slowly acting enzyme. The activity of rubisco increases markedly on exposure to light. How do the light reactions modify the activity of rubisco? Recall that CO2 must be added to lysine 201 of rubisco to form the carbamate that is essential for catalytic activity, a process facilitated by rubisco activase. Carbamate formation is favored by alkaline pH and high concentrations of Mg2+ ion in the stroma, both of which are consequences of the light-driven pumping of protons from the stroma into the thylakoid space. Magnesium ion concentration rises because Mg2+ ions from the thylakoid space are released into the stroma to compensate for the influx of protons. Thus, light leads to the generation of regulatory signals as well as ATP and NADPH.
Thioredoxin Plays a Key Role in Regulating the Calvin Cycle
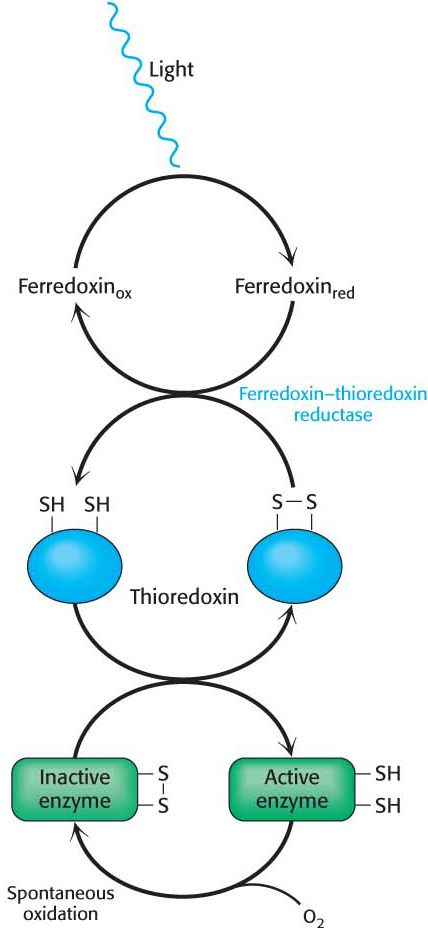
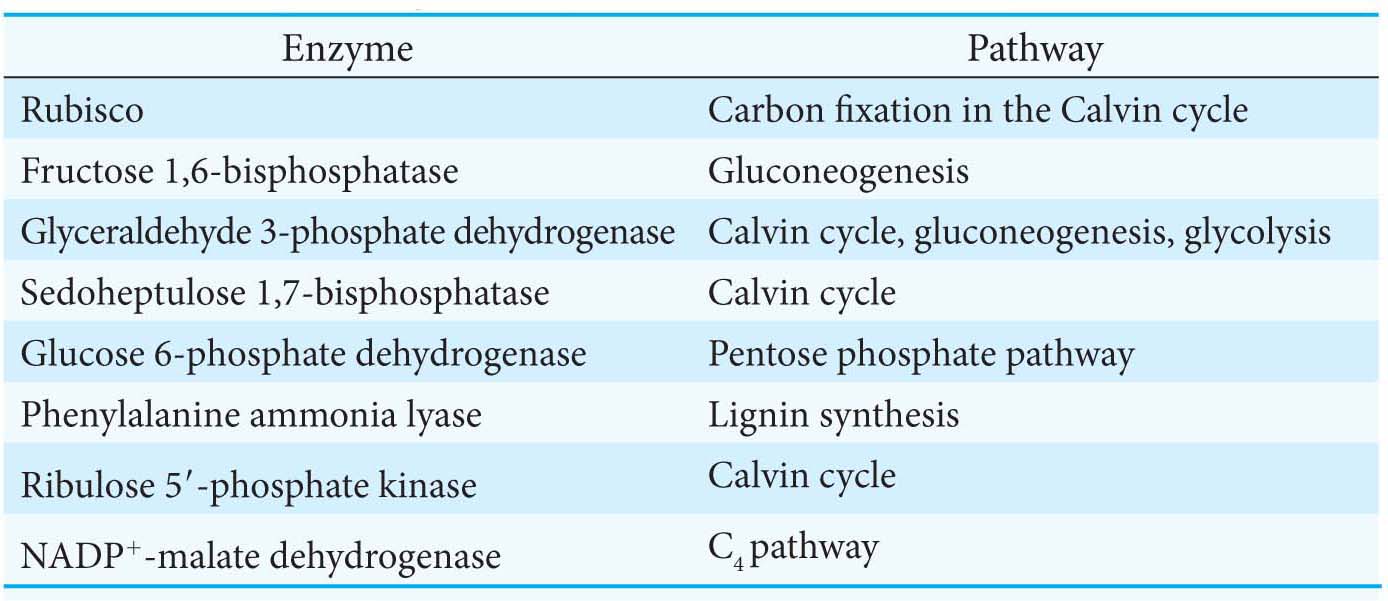
Light-driven reactions lead to electron transfer from water to ferredoxin and, eventually, to NADPH. Both reduced ferredoxin and NADPH regulate enzymes from the Calvin cycle. An important protein in these regulatory processes is thioredoxin, a 12-kDa protein containing neighboring cysteine residues. Thioredoxin cycles between an oxidized form with a disulfide bond between the two cysteines and a reduced sulfhydryl form (Figure 23.12). In chloroplasts, oxidized thioredoxin is reduced by ferredoxin in a reaction catalyzed by ferredoxin–thioredoxin reductase. The reduced form of thioredoxin activates rubisco and other Calvincycle enzymes, as well as many enzymes in other metabolic pathways in the cell by reducing disulfide bridges that control their activity including the chloroplast ATP synthase (Table 23.1). Thus, the activities of the light and dark reactions of photosynthesis are coordinated through electron transfer from reduced ferredoxin to thioredoxin and then to component enzymes containing regulatory disulfide bonds (Figure 23.13).
Figure 23.12 Thioredoxin. The oxidized form of thioredoxin contains a disulfide bond. When thioredoxin is reduced by reduced ferredoxin, the disulfide bond is converted into two free sulfhydryl groups. Reduced thioredoxin can cleave disulfide bonds in enzymes, activating certain Calvin-cycle enzymes and inactivating some degradative enzymes.
Figure 23.13 Enzyme activation by thioredoxin. Reduced thioredoxin activates certain Calvin-cycle enzymes by cleaving regulatory disulfide bonds.
Table 23.1 Enzymes regulated by thioredoxin
Page 436
Rubisco Also Catalyzes a Wasteful Oxygenase Reaction
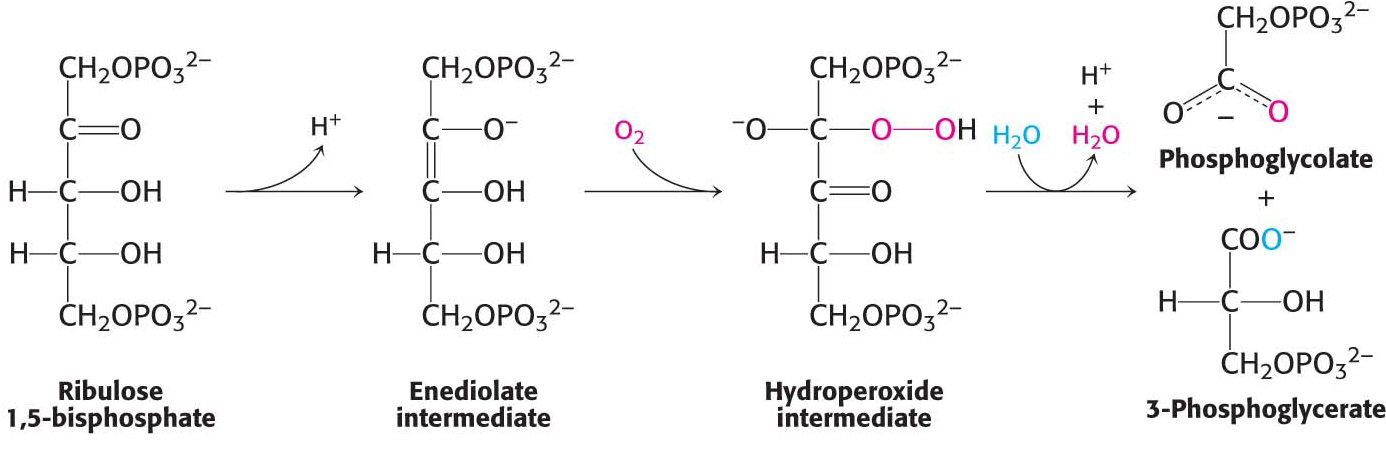
Rubisco is among the most important enzymes in life because it provides organic carbon for the biosphere. However, this enzyme has a wasteful side. Rubisco sometimes reacts with O2 instead of CO2, catalyzing a useless oxygenase reaction. The products of this reaction are phosphoglycolate and 3-phosphoglycerate (Figure 23.14). Phosphoglycolate is not a versatile metabolite. A salvage pathway recovers part of its carbon skeleton, recycling three of the four carbon atoms of two molecules of glycolate. However, one carbon atom is lost as CO2. This process is called photorespiration because, like cellular respiration, O2 is consumed and CO2 is released. However, photorespiration is wasteful because organic carbon is converted into CO2 without the production of ATP, NADPH, or another energy-rich metabolite. This wastefulness raises the question, what is the biochemical basis of this inefficiency? The results of structural studies show that, when the reactive enediolate intermediate is formed, loops close over the active site to protect the enediolate. A channel to the environment is maintained to allow access by CO2. However, like CO2, O2 is a linear molecule that also fits the channel. In essence, the problem lies not with the enzyme but in the unremarkable structure of CO2. Carbon dioxide lacks chemical features that would allow discrimination between it and other gases such as O2, and thus the oxygenase activity is an inevitable failing of the enzyme. Although evolutionary processes have enhanced the preference of rubisco for carboxylation—for instance, the rubisco of higher plants is eightfold as specific for carboxylation as that of photosynthetic bacteria—photorespiration still accounts for the loss of up to 25% of the carbon fixed. Another possibility exists, however. The oxygenase activity may not be an imperfection of the enzyme, but rather an imperfection in our understanding. Perhaps the oxygenase activity performs a biochemically important role that we have not yet discovered.
Figure 23.14 A wasteful side reaction. The reactive enediolate intermediate on rubisco also reacts with molecular oxygen to form a hydroperoxide intermediate, which then proceeds to form one molecule of 3-phosphoglycerate and one molecule of phosphoglycolate.
The oxygenase activity increases more rapidly with temperature than the carboxylase activity, presenting a problem for tropical plants. How do plants, such as sugarcane and corn that grow in hot climates, prevent very high rates of photorespiration? The solution to the problem illustrates one means by which photosynthesis responds to environmental conditions.
The C4 Pathway of Tropical Plants Accelerates Photosynthesis by Concentrating Carbon Dioxide
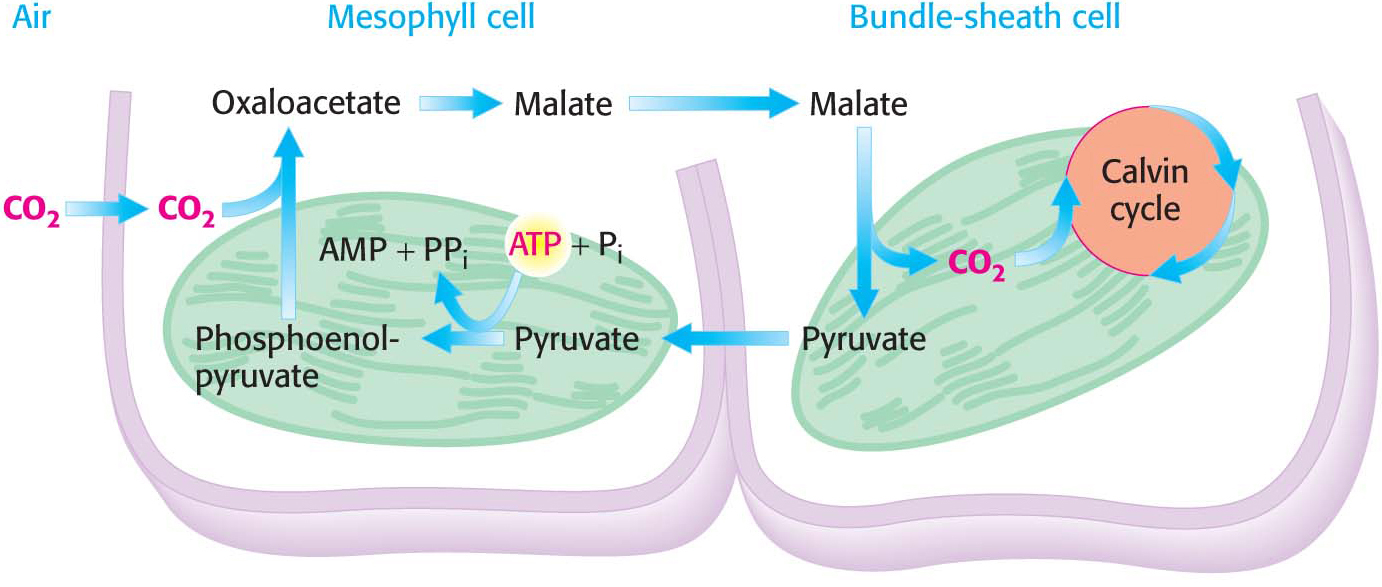
One means of overcoming the inherent oxygenase activity of rubisco is generating a high local concentration of CO2 at the site of the Calvin cycle in the photosynthetic cells. The key to accomplishing this is the synthesis of four-carbon (C4) compounds such as oxaloacetate that carry CO2 from mesophyll cells on the surfaces of leaves to interior bundle-sheath cells, which are the major sites of photosynthesis (Figure 23.15). Decarboxylation of the four-carbon compound in a bundle-sheath cell maintains a high concentration of CO2 at the site of the Calvin cycle. The resulting three-carbon compound, pyruvate, returns to the mesophyll cell for another round of carboxylation.
Figure 23.15 The C4 pathway. Carbon dioxide is concentrated in bundle-sheath cells by the expenditure of ATP in mesophyll cells.
Page 437
The C4pathway for the transport of CO2, also called the Hatch–Slack pathway after its discoverers Marshall Davidson Hatch and Charles Roger Slack, starts in a mesophyll cell with the condensation of CO2 and phosphoenolpyruvate to form oxaloacetate, in a reaction catalyzed by phosphoenolpyruvate carboxylase. Oxaloacetate is then converted into malate by an NADP+-linked malate dehydrogenase. Malate enters the bundle-sheath cell and is decarboxylated within the chloroplasts by a different isozymic form of NADP+-linked malate dehydrogenase. The released CO2 enters the Calvin cycle in the usual way by condensing with ribulose 1,5-bisphosphate. Pyruvate formed in this decarboxylation reaction returns to the mesophyll cell. Finally, phosphoenolpyruvate is formed from pyruvate by pyruvate-Pi dikinase. The net reaction of this C4 pathway is
Thus, the energetic equivalent of two ATP molecules is consumed in transporting CO2to the chloroplasts of the bundle-sheath cells. In essence, this process is active transport: the pumping of CO2 into the bundle-sheath cell is driven by the hydrolysis of one molecule of ATP to one molecule of AMP and two molecules of orthophosphate. The CO2 concentration can be 20-fold greater in the bundle-sheath cells than in the mesophyll cells as a result of this transport.
When the C4 pathway and the Calvin cycle operate together, the net reaction is
Note that 30 molecules of ATP are consumed per hexose molecule formed when the C4 pathway delivers CO2 to the Calvin cycle, in contrast with 18 molecules of ATP per hexose molecule in the absence of the C4 pathway. The elevated CO2 concentration resulting from the expenditure of an extra molecule of ATP enhances the rate of photosynthesis in tropical plants where light is abundant and minimizes the energy loss caused by photorespiration. Plants that rely on the C4 pathway are called C4plants.
?
QUICK QUIZ 2
Why is the C4 pathway valuable for tropical plants?
The C4 pathway allows the CO2 concentration to increase at the site of carbon fixation. High concentrations of CO2 inhibit the oxygenase reaction of rubisco. This inhibition is important for tropical plants because the oxygenase activity increases more rapidly with temperature than does the carboxylase activity.
Tropical plants with a C4pathway do little photorespiration because the high concentration of CO2in their bundle-sheath cells accelerates the carboxylase reaction relative to the oxygenase reaction. The geographical distribution of plants having this pathway (C4 plants) and those lacking it (C3 plants) can now be understood in molecular terms. C4plants have the advantage in a hot environment and under high illumination, which accounts for their prevalence in the tropics. C3plants, which consume only 18 molecules of ATP per hexose molecule formed in the absence of photorespiration (compared with 30 molecules of ATP for C4 plants), are more efficient than C4 plants in temperate environments (Figure 23.16).
Figure 23.16 C3 and C4 plants. (A) C3 plants, such as trees, account for 95% of plant species.(B) Corn is a C4 plant of tremendous agricultural importance.
Page 438
Crassulacean Acid Metabolism Permits Growth in Arid Ecosystems
Figure 23.17 Desert plants. Because of crassulacean acid metabolism, cacti are well suited to life in the desert.
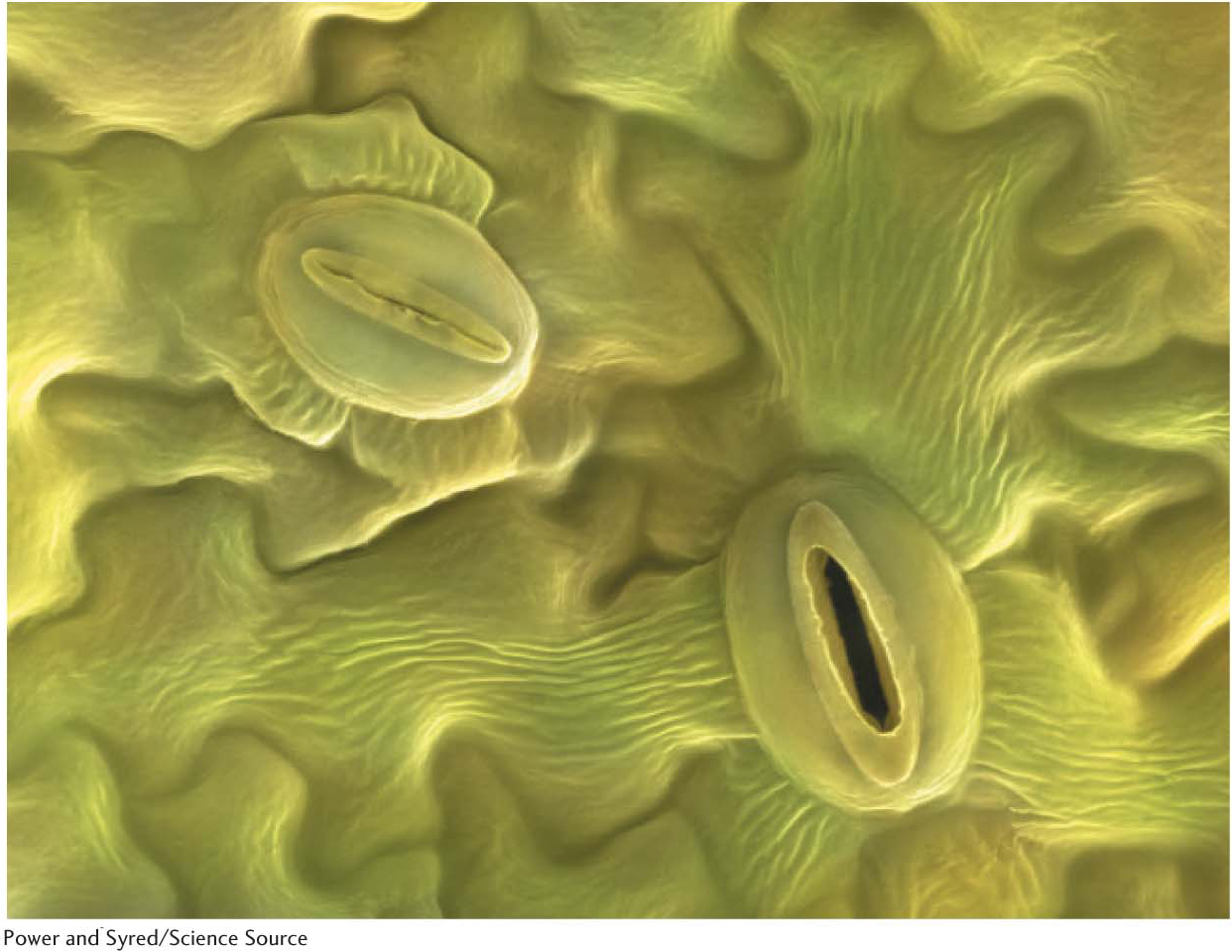
Figure 23.18 Stomata. An electron micrograph of an open stoma and a closed stoma.
Water is an important player in photosynthesis; if there is no water to provide electrons, oxygenic photosynthesis stops. Water availability is not a problem in temperate areas or hot, wet tropics, but it is not abundant in deserts (Figure 23.17). A crucial adaptation for such plants, and many non-desert plants as well, is the controlled opening and closing of the stomata (singular, stoma), microscopic pores located on the underside of plant leaves that are used for gas exchange. Such plants keep the stomata of their leaves closed in the heat of the day to prevent water loss (Figure 23.18). As a consequence, CO2 cannot be absorbed during the daylight hours when it is needed for hexose synthesis. Rather, CO2 enters the leaf when the stomata open at the cooler temperatures of night. To store the CO2 until it can be used during the day, such plants make use of an adaptation called crassulacean acid metabolism (CAM), named after the family Crassulaceae (including many succulents). Carbon dioxide is fixed by the C4 pathway into malate, which is stored in vacuoles. During the day, malate is decarboxylated and the CO2 becomes available to the Calvin cycle. In contrast with C4 plants, CAM plants separate CO2 accumulation from CO2 utilization temporally rather than spatially.
Although CAM plants do prevent water loss, the use of malate as the sole source of carbon dioxide comes at a metabolic cost. Because the malate storage is limited, CAM plants cannot generate CO2 as rapidly as it can be imported by C3 and C4 plants. Consequently, the growth rate of CAM plants is slower that that of C3 and C4 plants. The saguaros cactus, which can live up to 200 years and reach a height of 60 feet, can take 15 years to grow to only one foot in height.

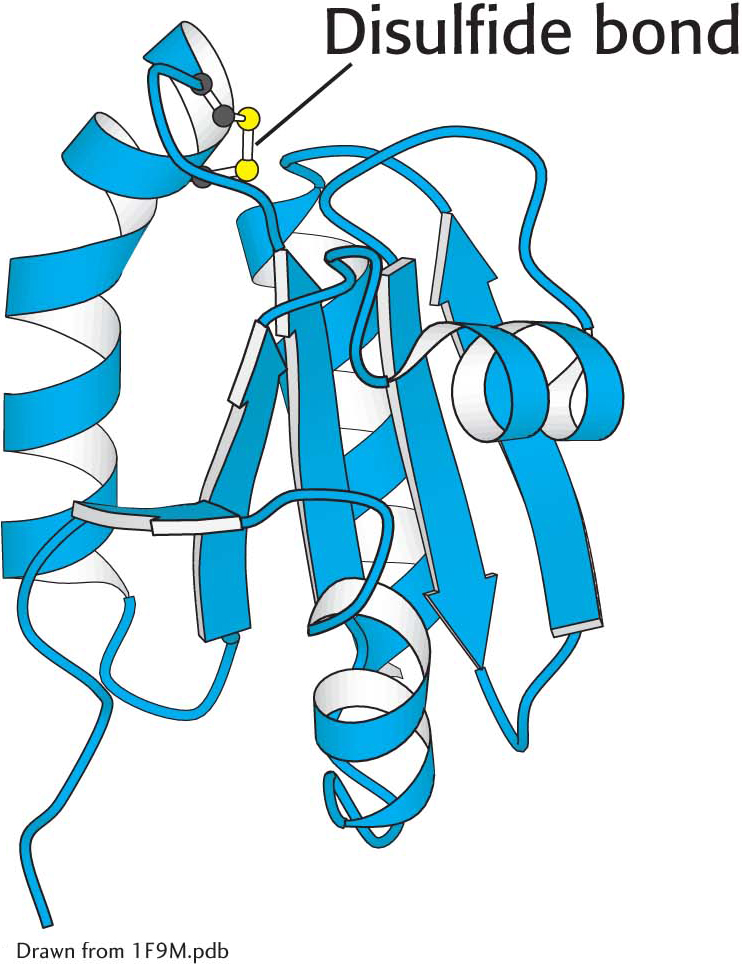
 Thioredoxin. The oxidized form of thioredoxin contains a disulfide bond. When thioredoxin is reduced by reduced ferredoxin, the disulfide bond is converted into two free sulfhydryl groups. Reduced thioredoxin can cleave disulfide bonds in enzymes, activating certain Calvin-
Thioredoxin. The oxidized form of thioredoxin contains a disulfide bond. When thioredoxin is reduced by reduced ferredoxin, the disulfide bond is converted into two free sulfhydryl groups. Reduced thioredoxin can cleave disulfide bonds in enzymes, activating certain Calvin-