
9.3 Hemoglobin Binds Oxygen Cooperatively
Like all allosteric proteins, hemoglobin displays quaternary structure. Human hemoglobin A, present in adults, consists of four subunits: two α subunits and two β subunits. The α and β subunits have similar three-
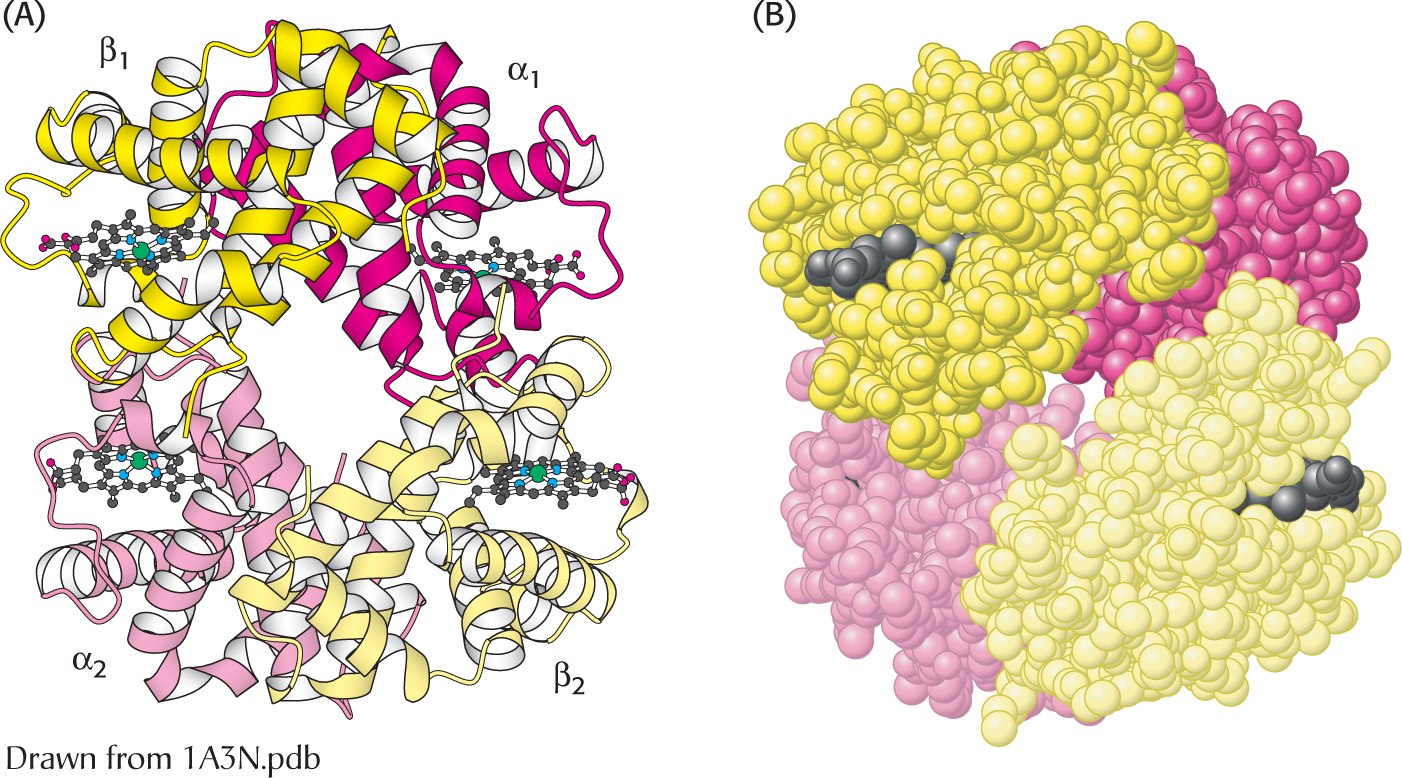
 The quaternary structure of deoxyhemoglobin. Hemoglobin, which is composed of two α chains and two β chains, functions as a pair of αβ dimers. (A) A ribbon diagram. (B) A space-
The quaternary structure of deoxyhemoglobin. Hemoglobin, which is composed of two α chains and two β chains, functions as a pair of αβ dimers. (A) A ribbon diagram. (B) A space-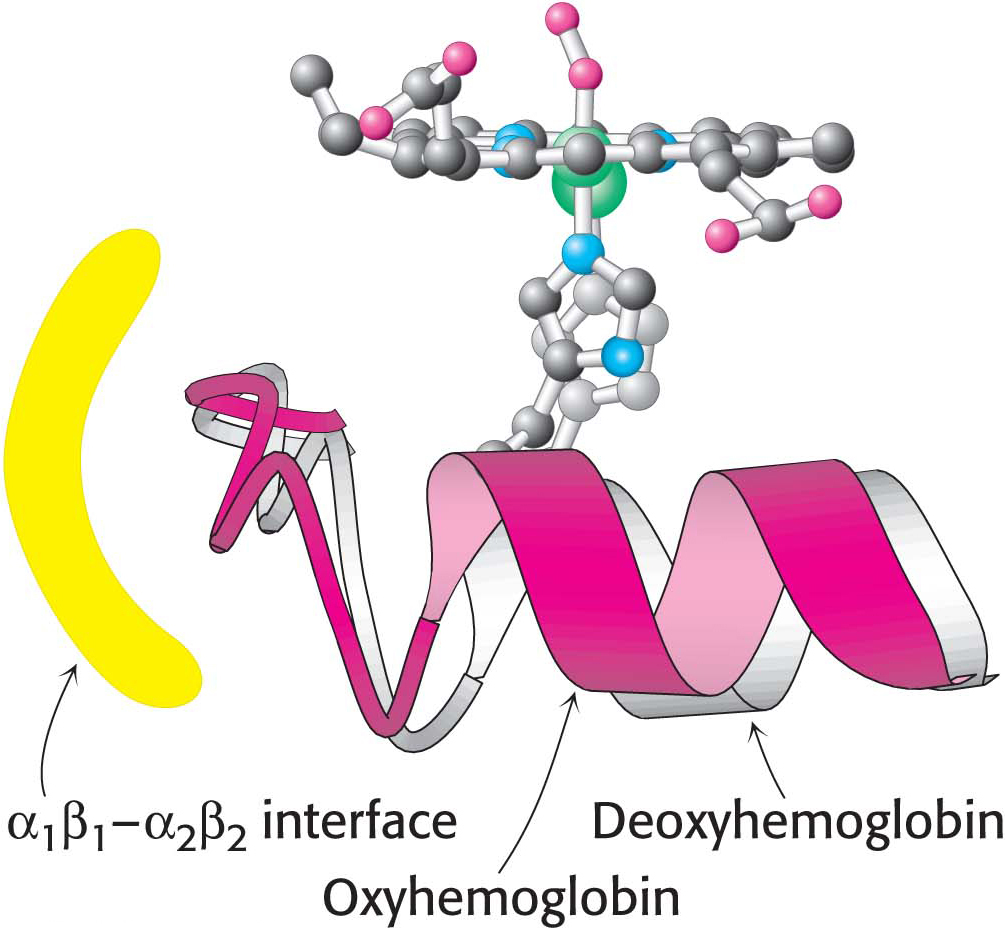
How does oxygen binding lead to the structural transition from the T state to the R state? When the iron ion moves into the plane of the porphyrin, the histidine residue bound in the fifth coordination site moves with it. This histidine residue is part of an α helix, which also moves (Figure 9.7). The carboxyl terminal end of this α helix lies in the interface between the two αβ dimers. Consequently, the structural transition at the iron ion is directly transmitted to the other subunits, resulting in substantial changes in quaternary structure that correspond to the T-
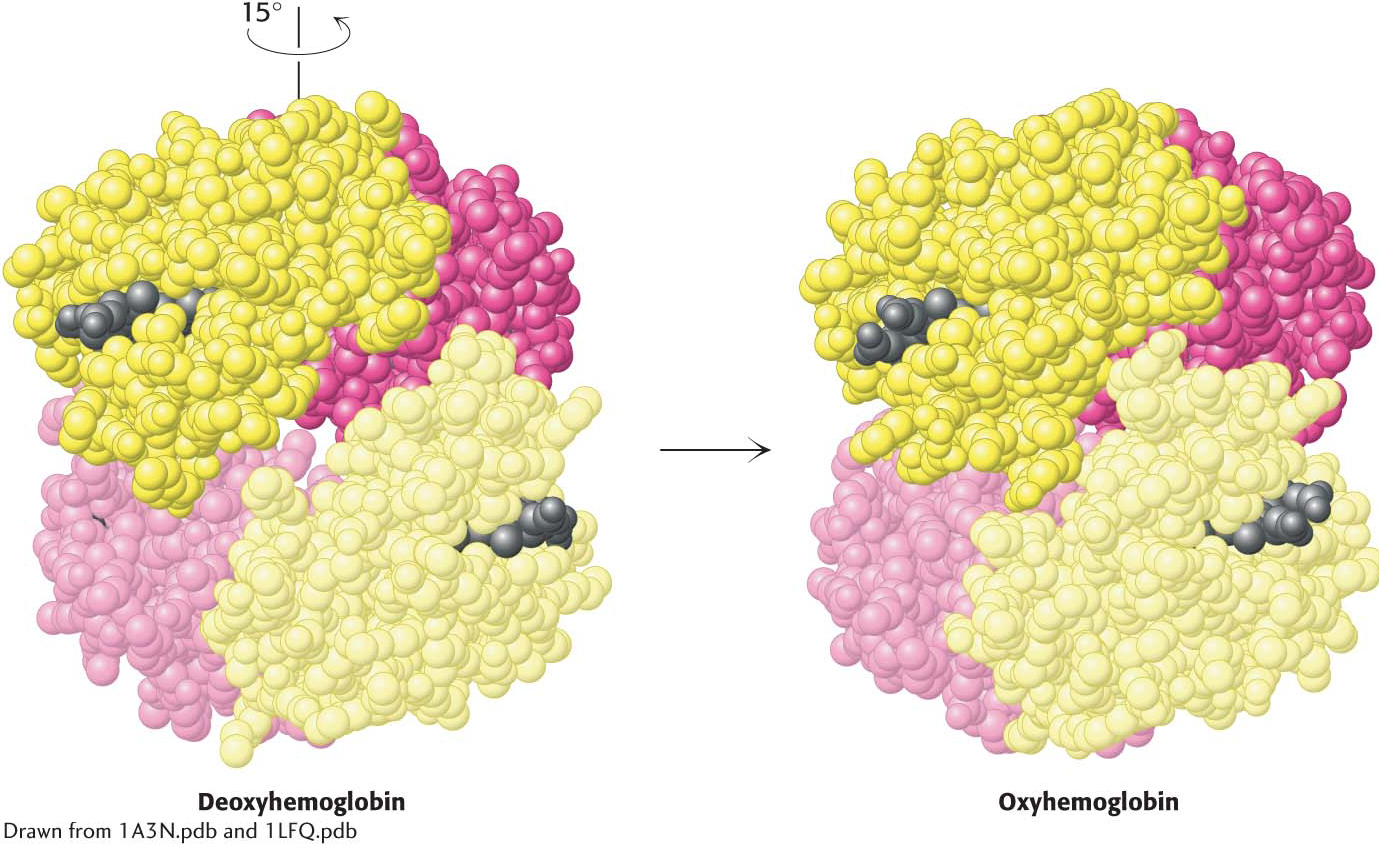 Quaternary structural changes on oxygen binding by hemoglobin. Notice that, on oxygenation, one αβ dimer shifts with respect to the other by a rotation of 15 degrees.
Quaternary structural changes on oxygen binding by hemoglobin. Notice that, on oxygenation, one αβ dimer shifts with respect to the other by a rotation of 15 degrees.
Recall that we considered two models for cooperative binding (Section 7.3). Is the cooperative binding of oxygen by hemoglobin best described by the concerted or the sequential model? Neither model in its pure form fully accounts for the behavior of hemoglobin. Instead, a combined model is required. Hemoglobin behavior is concerted in that hemoglobin with three sites occupied by oxygen is almost always in the quaternary structure associated with the R state. The remaining open binding site has an affinity for oxygen more than 20-