Chapter 2. DIVERSITY IV—FUNGI
Learning Objectives
General Purpose
Conceptual
- Be able to compare and contrast the major groups within the kingdom Fungi.
- Be able to recognize and describe the structures important in fungi reproduction.
Background Information
The kingdom Fungi is part of domain Eukarya and consists of structurally and nutritionally distinct organisms. Although fungi were once thought to be related to plants, recent molecular evidence suggests that fungi and animals diverged from a common ancestor (see Figure 12-1).
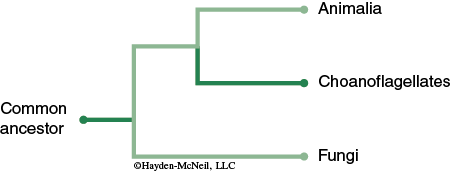
The organisms of the fungal lineage include mushrooms, rusts, smuts, puffballs, truffles, morels, molds, and yeasts, as well as many less well-known organisms (Alexopoulos et al., 1996). More than 70,000 species of fungi have been described; however, some estimates of total numbers suggest that 1.5 million species may exist (Hawksworth, 1991; Hawksworth et al., 1995).
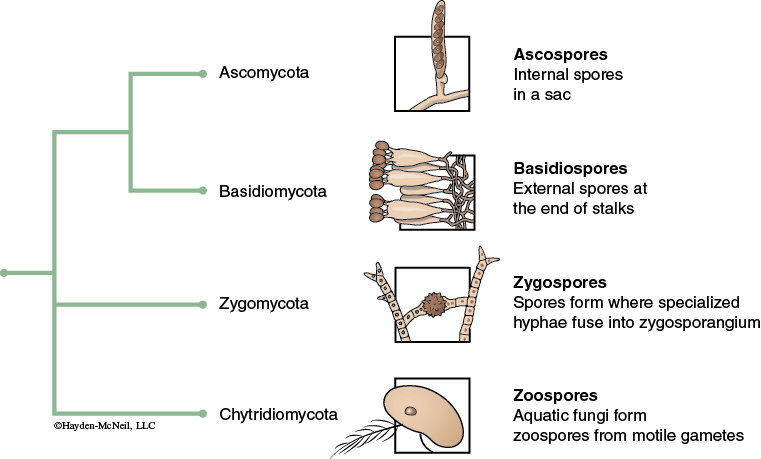
The various phyla of the Fungal kingdom can be defined in part by their sexual and asexual reproductive structures. These reproductive structures are not the macroscopic structures such as toadstools, but rather the microscopic structures associated with the spores produced during reproduction. Spores are presented in differing anatomical structures, e.g. ascospores of the Ascomycota phylum are contained within a sac-like ascus structure, while basidospores of the basidiomycota phylum are presented on the protruding surfaces of the basidia. Although molecular biology techniques sometimes supersede the morphological designation of fungal phyla, the reproductive structures inform the relationship between differing species (see Figure 12-2).
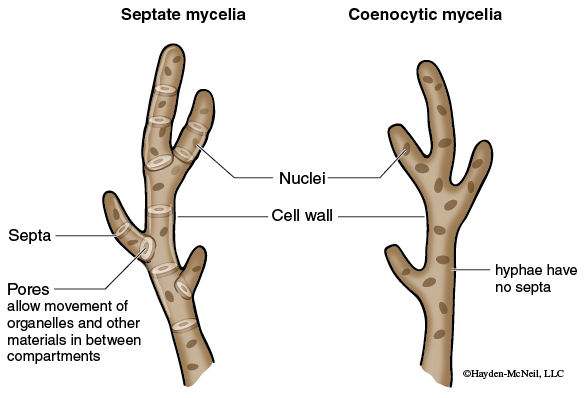
In general fungi are multicellular organisms composed of masses of filaments however there are a few species, including yeasts, which are unicellular. Individual filaments are called hyphae. The feeding network for the fungus is a mass of hyphae is called a mycelium. In most cases the mycelium is hidden within the food source. Growth of the hyphae occurs at the tip and can be very rapid. Depending on the species, hyphae are partitioned into cells by cross-walls called septa, or they may be aseptate or coenocytic (meaning multinucleate). The septa are usually perforated which allows the protoplast to flow from cell to cell (see Figure 12-3).
The cell walls (i.e., the hyphal walls) are composed primarily of chitin, a polysaccharide similar to the chitin found in the exoskeletons of insects.
Most biologists have seen dense filamentous fungal colonies growing on rich nutrient agar plates, but in nature the filaments can be much longer and the colonies less dense. When one of the filaments contacts a food supply, the entire colony mobilizes and reallocates resources to exploit the new food. Should all food become depleted, sporulation is triggered. Although the fungal filaments and spores are microscopic, the colony can be very large with individuals of some species rivaling the mass of the largest animals or plants.
Like bacteria, fungi are everywhere on Earth. Fungi reproduce using spores which are easily airborne and as such are all over the Earth. Fungi are eukaryotic heterotrophs that absorb their food by secreting hydrolytic digestive enzymes into a food source. These enzymes catalyze the breakdown of complex molecules into smaller molecules (i.e., polymers to monomers) that can be absorbed into the fungal cells. Rather than requiring a stomach to accomplish digestion, fungi live in their own food supply and simply grow into new food as the local environment becomes nutrient depleted. Because of this absorptive nutrition fungi must normally live in environments that are rich organic substances.
This absorptive mode of nutrition allows specialization of fungi as 1) decomposers (saprobes), 2) parasites, or 3) mutualistic symbionts.
Saprobic fungi are extremely beneficial because the digestive enzymes they secrete are responsible for decomposing dead plants and animals, fallen leaves, and wastes of living organisms, making it possible to recycle chemical elements. Within their varied natural habitats fungi usually are the primary decomposer organisms present. Many species are free-living saprobes (users of carbon fixed by other organisims) in woody substrates, soils, leaf litter, dead animals, and animal exudates. The larger cavities eaten out of living trees by wood-decaying fungi provide nest holes for a variety of animals. Some fungi actually are very specific, decomposing organic material from burned soil or animal urine. Other saprobic fungi secrete enzymes that digest materials valuable to us, causing wood to rot (see Figure 12-4), fabrics to mildew, and mold to grow on food.

On the LSU campus there are a number of fungi that specialize on living on the bark of live oak trees. The reproductive fruiting bodies on these can be seen most of the year on the trunks and undersides of branches (see Figure 12-5).

In some low nitrogen environments several independent groups of fungi have adaptations such as nooses and sticky knobs with which to trap and degrade nematodes and other small animals. A number of references on fungal ecology are available (Carroll and Wicklow, 1992; Cooke and Whipps, 1993; Dix and Webster, 1995).
Parasitic fungi secrete enzymes that digest nutrients from the cells of living hosts. They cause many plant diseases, such as leaf spots, rust, smuts, and many ascomycetes such as the agents of Dutch elm disease and chestnut blight. Among the other well-known associations are fungal parasites of animals which cause a number of diseases in humans including ringworm, athlete’s foot, vaginal infections and thrush. Individuals with compromised immune systems are particularly vulnerable to fungal infections and may succumb to diseases caused by Pneumocystis (a type of pneumonia that affects individuals with suppressed immune systems), Coccidioides (valley fever), Ajellomyces (blastomycosis and histoplasmosis), and Cryptococcus (cryptococcosis) (Kwon-Chung and Bennett, 1992).
Mutualistic fungi are those that live in mutually beneficial symbiotic relationships. Two common mutualistic fungal symbiotic relationships are lichens and mycorrhizae. Lichens are symbiotic associations of unicellular photosynthetic organisms (either algae or bacteria) and fungi. These unique partnerships can grow in places that other organisms cannot grow (see Figure 12-6).

Although lichens may not be seen frequently in polluted cities, they can form the dominant growth in Nordic environments.
Mycorrhizae are symbiotic associations of fungi with the roots of vascular plants (see Figure 12-7). Nearly every vascular plant growing in the wild has a mycorrhizal relationship with a fungus.
Leaf and stem endophytes are a more recent discovery of mutualistic fungal relationship. Some of the fungi in these relationships can protect the plants they inhabit from herbivory and even influence flowering and other aspects of plant reproductive biology.
Fungi reproduce by the formation of spores. Spores may be produced asexually and sexually depending on the species. The specialized spore-producing structure forms on hyphae (see Figure 12-8).
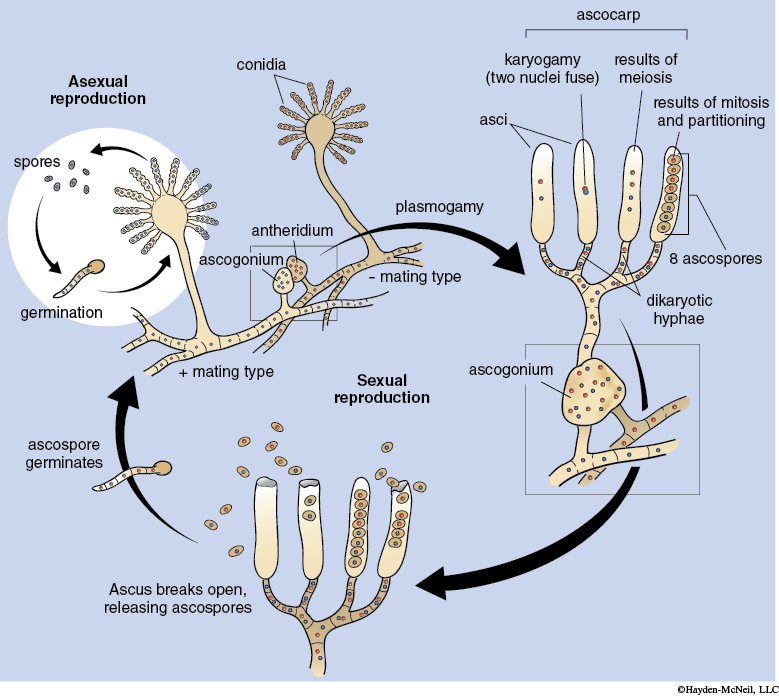
Spores are produced in the reproductive fruiting bodies. Many species have elaborate and sometimes bizarre types of fruiting bodies to enclose, protect, and promote the spreading fungal spores to new habitats. Spores are usually dispersed by wind or water (including rain), or by animals. The spores will germinate to form a new fungus when conditions are favorable. Depending on the species spore formation may occur as part of sexual or asexual reproduction. In addition, a few fungi regularly reproduce asexually by fragmentation of their hyphae. Other species can also use this method as an alternative means for reproduction and dispersal.
Sexual reproduction in many fungal species includes two distinct stages. In the first stage, plasmogamy, the cytoplasm of two compatible hyphae or asexual spores fuse resulting in dikaryotic hyphae designated as n + n because the haploid nuclei do not fuse. During the second stage, karyogamy, the haploid nuclei fuse, resulting in the diploid stage. Some fungi may remain dikaryotic for years before the diploid stage is formed. The diploid stage undergoes meiosis to produce haploid spores.
Many fungi are economically useful. Commercial uses of fungi and their enzymes include the production of wine, beer, certain cheeses, various antibiotics and bread. Other fungi, such as mushrooms, are eaten regularly by humans.
For more about the general characteristics of fungal organisms view the following video: