Chapter 2. EVOLUTION II—SELECTION
Learning Objectives
General Purpose
Conceptual
- Gain an understanding of how the term adaption can be used in biology and how this term applies to evolution.
- Define the term gene pool and explain how it relates to the concept of evolution.
- Gain an understanding of the concept of genetic equilibrium and the conditions required for it to occur.
- Gain an understanding of the relationship between genetic equilibrium and evolution.
- Gain an understanding of the Hardy-Weinberg equation and how it relates to population evolution.
- Gain an understanding of the impact of selection, mutation, and population size.
Procedural
- Be able to use the Hardy-Weinberg equation to calculate allele frequencies and the percentages of genotypes in a population.
Recall that the Hardy-Weinberg equation can be used to calculate the genotype frequencies of a pair of alleles at a gene locus in an established population. It can also be used to predict genotype frequencies of the next generation as long as there have been no changes in the gene pool. When there have been no changes, then genetic equilibrium occurs and the population does not evolve with respect to those alleles.
Genetic equilibrium will only occur when
- The population is large enough to be unaffected by random gene changes.
- There is no gene flow (immigration or emigration).
- There are no mutations occurring or there is mutational equilibrium.
- There is random mating.
- There is no natural selection.
Small Population
When populations are small enough, random events that impact the population will have substantial effect on the gene pool. Notice that the terms here are relative. If the population is 50 individuals, an event that kills off or simply prevents the reproduction of 5 individuals may be a large impact on the gene pool.
Small populations are also more prone to genetic drift. Genetic drift is a change in gene pool that occurs purely as a result of chance. Genetic drift always influences gene frequencies to some degree, no matter how large the population. In small populations of less than approximately 100 individuals, genetic drift may be strong enough to significantly alter the allele frequencies.
In the most extreme case, genetic drift leads to the loss of certain alleles and the fixation of another. Gene fixation occurs when only one allele for a gene is present in a population. Fixation results in a lack of variability for a particular gene in that population.
It has been observed that genetic drift is particularly evident after a new population is started by only a few individuals. Imagine that by chance a small number of individuals found a colony and then by chance gamete formation, only certain of the founders’ alleles are passed on to the next generation. As a result, a severe reduction in genetic variation will take place and genetic drift will occur (see Figure 2-1).
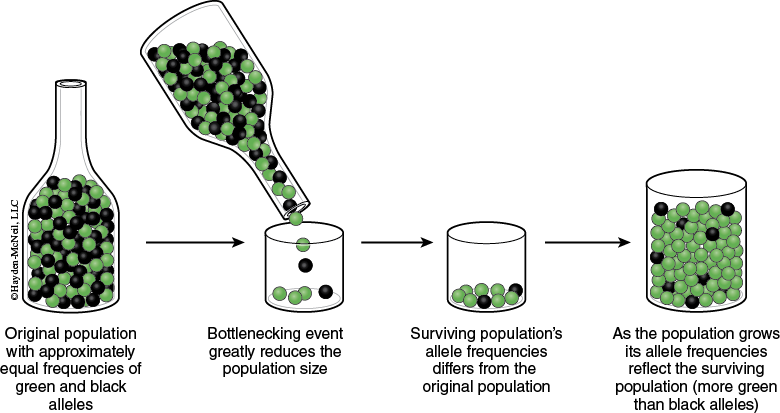
Nonrandom Mating
In populations that use sexual reproduction, there is a potential for the mating to be influenced by a number of factors. If in fact any form of influence occurs in the mating process, then it is likely that the mating will be nonrandom. In the following figure a case of nonrandom mating in a bird species is examined (see Figure 2-2).
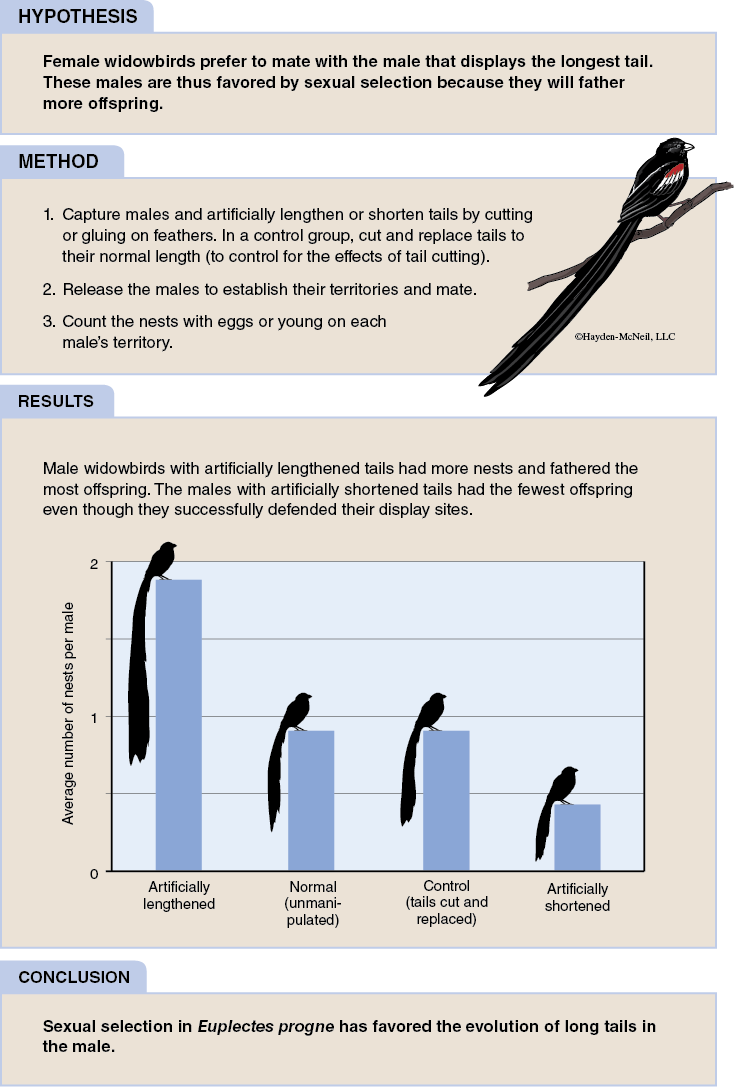
Selection
The process of selection can be a natural one (survival of the fittest) in which individuals of a certain phenotype (or genotype) are able to reproduce more frequently than individuals that have a less favored phenotype (or genotype). This difference in reproduction can be the result of differences in survival (can’t reproduce if you’re dead) or the result in differences in reproduction rates. The case of nonrandom mating would result in selection.
Selection based on survival of the fittest is sometimes referred to as adaptation. This term is also used many other ways in biology, so it is important to understand the differences in the way this term can be applied to the different scenarios, and it is especially critical to understand what it means in relation to evolution.
Please view the following video for a description of some of the ways the term adaptation is used in biology.
Selection can also be artificial. This is the case with any form of selective breeding from crop development to animal breeding. In these cases, only the individuals with the desirable traits are selected for reproduction.
Heterozygous Advantage
It is frequently the case where selection or nonrandom mating occurs to favor one phenotype over another. In the case of traditional Mendelian inheritance this would mean either the heterozygous or homozygous dominant phenotype would have differential success compared to the homozygous recessive. However, in the case of non-Mendelian inheritance (incomplete dominance, codominance etc.) different scenarios can occur. For example, what happens if both of the homozygous conditions are selected against? A real-world example of this is malaria, a disease caused by a parasitic protozoan. This disease can occur in many tropical and subtropical ecosystems. Malaria can be fatal; however when malaria is found in areas which also have a high frequency of the sickle cell anemia trait an interesting selection occurs.
What became apparent after years of investigation was that heterozygous individuals (Aa) for the sickle cell anemia trait did not die of malaria. The sickle cell hemoglobin trait was not expressed in them sufficiently to cause sickle cell anemia. The presence of the sickle cell gene was sufficient to make heterozygous individuals undesirable hosts for malaria parasites. The heterozygous individuals suffer neither fatal sickle cell anemia nor fatal malaria. Thus the heterozygous individuals have an advantage in these specific circumstances. The individuals who are homozygous for the normal hemoglobin gene are more likely to contract malaria, whereas the individuals that are homozygous for the sickle cell anemia trait develop sickle cell anemia.
Pre-Lab Quiz
Proceed to the Pre-Lab Quiz