Chapter 2. EVOLUTION III—OVERVIEW
Learning Objectives
General Purpose
Conceptual
- Gain an understanding of the impact of selection.
- Gain an understanding of how and why replica plating can be used to test a population against different selective pressures.
The development of resistance of an organism to certain chemicals is a case of artificial selection-induced evolution. This is true regardless of the nature of the chemicals being used (e.g., herbicides, fungicides, pesticides, antibiotics, etc.). The resistance arises naturally via the process of mutation. In a population under no selective pressure from the presence of a chemical treatment, there is no survival advantage conferred by these mutations. In those cases the mutated gene may disappear from the population simply from random chance (i.e., genetic drift). However, in cases where there is survival pressure from the presence of a chemical in the environment, then the mutation can confer a survival advantage. If those individuals with the mutated gene survive better, then they have an increased fitness which should lead to a change in the genetic makeup of the population. That’s evolution.
There are many examples of resistance developing in populations. One such example is warfarin resistance in the Norweigan brown rat, Rattus norvegicus (see Figure 3-1).
Europe did not have Norwegian brown rats, Rattus norvegicus, until the eighteenth century (Sullivan, 2004). Prior to this time, black rats were the prominent vermin in Europe. The brown rat was believed to have originated in China and spread on ships wherever men traveled. The rat was mistakenly believed to have come to England on ships from Norway (hence the name), although the source was probably Denmark. It was most likely introduced to the United States of America during the late 1700s. R. norvegicus has been incredibly successful, displacing the black rat as the dominant rat around the world. If you see a rat in the wild, most likely it is of the species R. norvegicus.
R. norvegicus live in family groups and have a limited range of about 700 m2 (Timm, 2005). Although they rarely stray far from their home, if a population becomes overcrowded, rats will migrate in large groups. Rats will start to breed at 11 weeks and have an average litter size of about 7. Rats can live for two years (Calhoun, 1963); however, in many environments, the yearly mortality rate will be up to 99% (Drummond, 1966). When the population is sparse, litter size increases and the population rapidly rebounds.
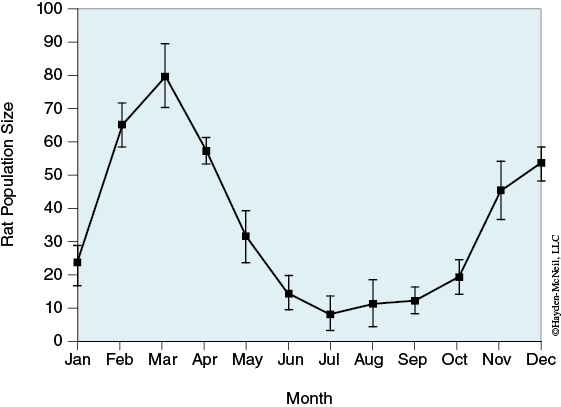
Rat populations are difficult to estimate and few reliable numbers exist. One extensive study found that Baltimore had an estimated population of 48,000 (Easterbrook, 2004). In addition to the difficulty of establishing a population estimate of rats at one point in time, rat populations can vary widely even in the same location (see Figure 3-2). Over the course of a year, studies have shown that population varied over tenfold (McGuire, 2006; Villa, 1997). This level of population variation can be attributed to reproduction and predation rather than migration.
It is estimated that rats produce $19 million in economic damage in the form of the destruction of food and property. In addition, rats may carry pathogenic organisms, which can cause diseases, including bubonic plague (fleas on rats are carriers), trichinosis, typhoid, and tuberculosis.
One way to control rat populations is by putting out poisoned bait. However, Norwegian rats are difficult to poison because they are initially cautious with new foods and then learn to avoid baits that quickly induce sickness. Warfarin was seen as a promising poison because death due to warfarin occurs days after ingestion, so rats would not link its effects with the poisoned bait. Warfarin was introduced as a rat poison in the 1950s and quickly became a popular method of control.
Death due to warfarin results from an inhibition of vitamin K reductase, an enzyme required for blood coagulation. Mutations in the vitamin K reductase gene can confer warfarin resistance. When this mutation is present in a rat population and warfarin is extensively used for rat poison, selection will favor the warfarin-resistant phenotype. Warfarin resistance was first noted in European rats in 1958 and has subsequently been found in many other parts of the world.
As its name implies, vitamin K reductase requires vitamin K to function. Different alleles at this locus vary in their vitamin requirements. One allele (designated R, resistant to warfarin) confers warfarin resistance, but also results in a twentyfold increase in vitamin K requirement over the other allele (designated S, susceptible to warfarin) for its primary physiological function (Pelz, 2007). In areas where vitamin K is readily available, the two allele types are equally fit. In areas where rats do not have sufficient vitamin K, homozygous RR individuals are less fit (see Table 3-1). However, since the R allele confers resistance to warfarin, rats expressing this allele are favored in areas where warfarin is used, despite its disadvantage in vitamin K utilization.
Table 3-1. Fitness of genotypes of R. norvegicus population with vitamin K reductase mutation requiring additional vitamin K for proper functioning in the presence or absence of warfarin. Fitness in low vitamin K availability estimated from Pelz (2007).

Pre-Lab Quiz
Proceed to the Pre-Lab Quiz