13.5 Gap Junctions Allow Ions and Small Molecules to Flow Between Communicating Cells
The ion channels that we have considered thus far have narrow pores and are moderately to highly selective in the ions that they allow to pass through them. They are closed in the resting state and have short lifetimes in the open state, typically a millisecond, that enable them to transmit frequent neural signals. We turn now to a channel with a very different role. Gap junctions, also known as cell-to-cell channels, serve as passageways between the interiors of contiguous cells. Gap junctions are clustered in discrete regions of the plasma membranes of apposed cells. Electron micrographs of sheets of gap junctions show them tightly packed in a regular hexagonal array (Figure 13.31). An approximately 20-Å central hole, the lumen of the channel, is prominent in each gap junction. These channels span the intervening space, or gap, between apposed cells (hence, the name “gap junction”). The width of the gap between the cytoplasms of the two cells is about 35 Å.
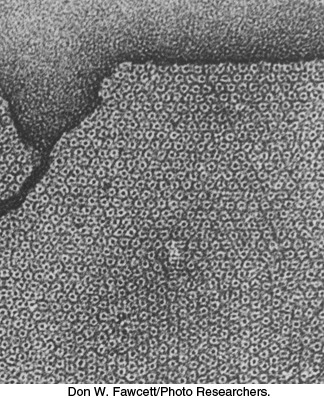
Figure 13.31: Gap junctions. This electron micrograph shows a sheet of isolated gap junctions. The cylindrical connexons form a hexagonal lattice having a unit-cell length of 85 Å. The densely stained central hole has a diameter of about 20 Å.
[Don W. Fawcett/Photo Researchers.]
Small hydrophilic molecules as well as ions can pass through gap junctions. The pore size of the junctions was determined by microinjecting a series of fluorescent molecules into cells and observing their passage into adjoining cells. All polar molecules with a mass of less than about 1 kDa can readily pass through these cell-to-cell channels. Thus, inorganic ions and most metabolites (e.g., sugars, amino acids, and nucleotides) can flow between the interiors of cells joined by gap junctions. In contrast, proteins, nucleic acids, and polysaccharides are too large to traverse these channels. Gap junctions are important for intercellular communication. Cells in some excitable tissues, such as heart muscle, are coupled by the rapid flow of ions through these junctions, which ensures a rapid and synchronous response to stimuli. Gap junctions are also essential for the nourishment of cells that are distant from blood vessels, as in lens and bone. Moreover, communicating channels are important in development and differentiation. For example, the quiescent uterus transforms to a forcefully contracting organ at the onset of labor; the formation of functional gap junctions at that time creates a syncytium of muscle cells that contract in synchrony.
A cell-to-cell channel is made of 12 molecules of connexin, one of a family of transmembrane proteins with molecular masses ranging from 30 to 42 kDa. Each connexin molecule contains four membrane-spanning helices (Figure 13.32A). Six connexin molecules are hexagonally arrayed to form a half-channel, called a connexon or hemichannel. Two connexons join end to end in the intercellular space to form a functional channel between the communicating cells (Figure 13.32B). Each connexon adopts a funnel-shape: at the cytoplasmic face, the inner diameter of the channel is 35 Å, while at its innermost point, the pore narrows to a diameter of 14 Å (Figure 13.32C). Cell-to-cell channels differ from other membrane channels in three respects: (1) they traverse two membranes rather than one; (2) they connect cytoplasm to cytoplasm, rather than to the extracellular space or the lumen of an organelle; and (3) the connexons forming a channel are synthesized by different cells. Gap junctions form readily when cells are brought together. A cell-to-cell channel, once formed, tends to stay open for seconds to minutes. They are closed by high concentrations of calcium ion and by low pH. The closing of gap junctions by Ca2+ and H+ serves to seal normal cells from injured or dying neighbors. Gap junctions are also controlled by membrane potential and by hormone-induced phosphorylation.
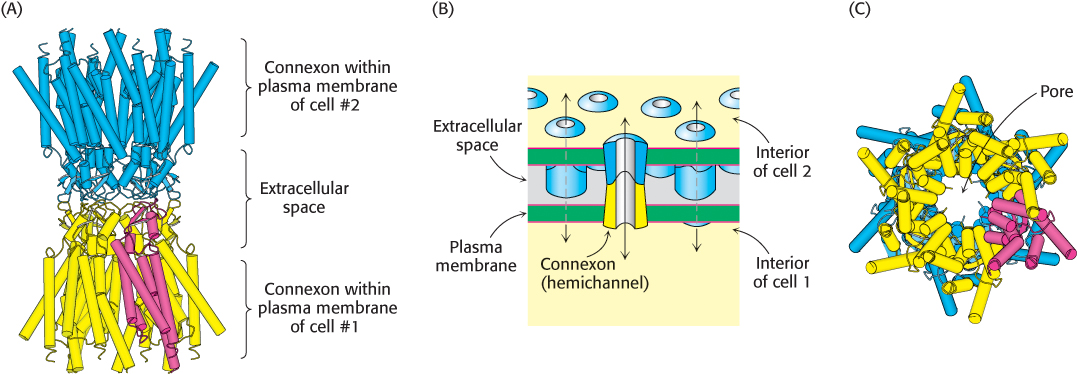
Figure 13.32: Structure of a gap junction. (A) Six connexins join to form a connexon, or hemichannel, within the plasma membrane (yellow). A single connexin monomer is highlighted in red. The extracellular region of one connexon binds to the same region of a connexon from another cell (blue), forming a complete gap junction. (B) Schematic view of the gap junction, oriented in the same direction as in (A). (C) A bottom-up view looking through the pore of a gap junction. This perspective is visualized in Figure 13.31. [(A) and (C) Drawn from 2ZW3.pdb; (B) Information from Dr. Werner Loewenstein.]
The human genome encodes 21 distinct connexins. Different members of this family are expressed in different tissues. For example, connexin 26 is expressed in key tissues in the ear. Mutations in this connexin are associated with hereditary deafness. The mechanistic basis for this deafness appears to be insufficient transport of ions or second-messenger molecules, such as inositol trisphosphate, between sensory cells.