33.1 A Wide Variety of Organic Compounds Are Detected by Olfaction
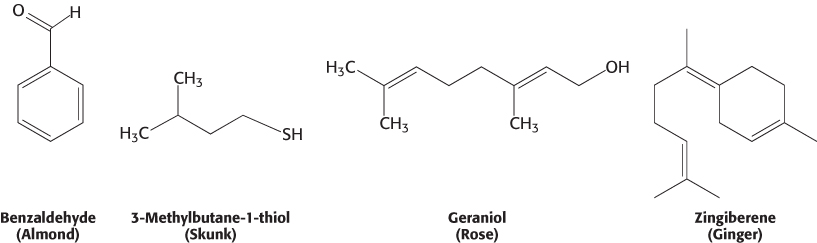
Human beings can detect and distinguish thousands of different compounds by smell, often with considerable sensitivity and specificity. Most odorants are small organic compounds with sufficient volatility that they can be carried as vapors into the nose. For example, a major component responsible for the odor of almonds is the simple aromatic compound benzaldehyde, whereas the sulfhydryl compound 3-methylbutane-1-thiol is a major component of the odor of skunks.
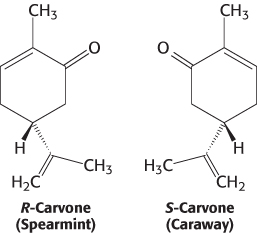
What properties of these molecules are responsible for their odors? First, the shape of the molecule rather than its other physical properties is crucial. We can most clearly see the importance of shape by comparing molecules such as those responsible for the odors of spearmint and caraway.
These compounds are identical in essentially all physical properties such as hydrophobicity because they are exact mirror images of one another. Thus, the odor produced by an odorant depends not on a physical property but on the compound’s interaction with a specific binding surface, most likely a protein receptor. Second, some human beings (and other animals) suffer from specific anosmias; that is, they are incapable of smelling specific compounds even though their olfactory systems are otherwise normal. Such anosmias are often inherited. These observations suggest that mutations in individual receptor genes lead to the loss of the ability to detect a small subset of compounds.
Olfaction is mediated by an enormous family of seven-transmembrane-helix receptors
Odorants are detected in a specific region of the nose, called the main olfactory epithelium, that lies at the top of the nasal cavity (Figure 33.2). Approximately 1 million sensory neurons line the surface of this region. Cilia containing the odorant-binding protein receptors project from these neurons into the mucous lining of the nasal cavity.
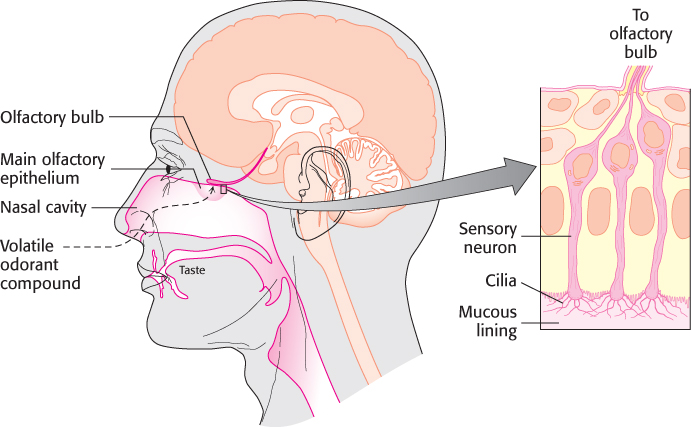
Figure 33.2: The main nasal epithelium. This region of the nose, which lies at the top of the nasal cavity, contains approximately 1 million sensory neurons. Nerve impulses generated by odorant molecules binding to receptors on the cilia travel from the sensory neurons to the olfactory bulb.
Biochemical studies in the late 1980s examined isolated cilia from rat olfactory epithelia that had been treated with odorants. Exposure to the odorants increased the cellular level of cyclic AMP, and this increase was observed only in the presence of GTP. On the basis of what was known about signal-transduction systems, the participation of cAMP and GTP strongly suggested the involvement of a G protein and, hence, 7TM receptors. Indeed, Randall Reed purified and cloned a G-protein α subunit, termed G(olf), which is uniquely expressed in olfactory cilia. The involvement of 7TM receptors suggested a strategy for identifying the olfactory receptors themselves. Complementary DNAs were sought that (1) were expressed primarily in the sensory neurons lining the nasal epithelium, (2) encoded members of the 7TM-receptor family, and (3) were present as a large and diverse family to account for the range of odorants. Through the use of these criteria, cDNAs for odorant receptors from rats were identified in 1991 by Richard Axel and Linda Buck.
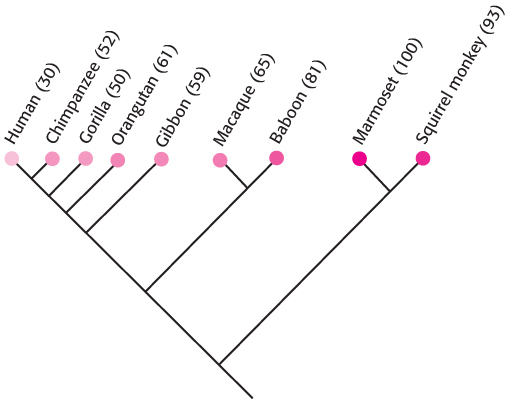
Figure 33.3: Evolution of odorant receptors. Odorant receptors appear to have lost function through conversion into pseudogenes in the course of primate evolution. The percentage of OR genes that appear to be functional for each species is given in parentheses.
 The odorant receptor (hereafter, OR) family is even larger than expected: more than 1000 OR genes are present in the mouse and the rat, whereas the human genome encodes approximately 350 ORs. In addition, the human genome includes approximately 500 OR pseudogenes containing mutations that prevent the generation of a full-length, proper odorant receptor. The OR family is thus one of the largest gene families in human beings. Further analysis of primate OR genes reveals that the fraction of pseudogenes is greater in species more closely related to human beings (Figure 33.3). Thus, we may have a glimpse at the evolutionary loss of acuity in the sense of smell as higher mammals presumably became less dependent on this sense for survival. For rodents that are highly dependent on their sense of smell, essentially all OR genes encode functional proteins.
The odorant receptor (hereafter, OR) family is even larger than expected: more than 1000 OR genes are present in the mouse and the rat, whereas the human genome encodes approximately 350 ORs. In addition, the human genome includes approximately 500 OR pseudogenes containing mutations that prevent the generation of a full-length, proper odorant receptor. The OR family is thus one of the largest gene families in human beings. Further analysis of primate OR genes reveals that the fraction of pseudogenes is greater in species more closely related to human beings (Figure 33.3). Thus, we may have a glimpse at the evolutionary loss of acuity in the sense of smell as higher mammals presumably became less dependent on this sense for survival. For rodents that are highly dependent on their sense of smell, essentially all OR genes encode functional proteins.
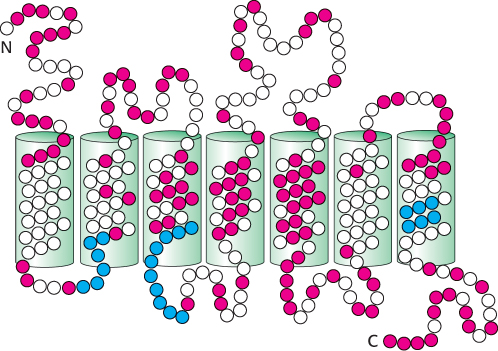
Figure 33.4: Conserved and variant regions in odorant receptors. Odorant receptors are members of the 7TM-receptor family. The green cylinders represent the seven presumed transmembrane helices. Strongly conserved residues characteristic of this protein family are shown in blue, whereas highly variable residues are shown in red.
The OR proteins are typically 20% identical in sequence with the β-adrenergic receptor (Section 14.1) and from 30% to 60% identical with one another. Several specific sequence features are present in most or all OR family members (Figure 33.4). The central region, particularly transmembrane helices 4 and 5, is highly variable, suggesting that this region is the site of odorant binding. That site must be different in odorant receptors that bind distinct odorant molecules.
What is the relation between OR gene expression and the individual neuron? Interestingly, each olfactory neuron expresses only a single OR gene, among hundreds available. Apparently, the precise OR gene expressed is determined largely at random. After one OR gene is expressed and a functional OR protein is produced, the expression of all other OR genes is suppressed by a feedback mechanism that remains to be fully elucidated.
The binding of an odorant to an OR on the neuronal surface initiates a signal-transduction cascade that results in an action potential (Figure 33.5). The ligand-bound OR activates G(olf), the specific G protein mentioned earlier. G(olf) is initially in its GDP-bound form. When activated, it releases GDP, binds GTP, and releases its associated βγ subunits. The α subunit then activates a specific adenylate cyclase, increasing the intracellular concentration of cAMP. The rise in the intracellular concentration of cAMP activates a nonspecific cation channel that allows calcium and other cations into the cell. The flow of cations through the channel depolarizes the neuronal membrane and initiates an action potential. This action potential, combined with those from other olfactory neurons, leads to the perception of a specific odor.
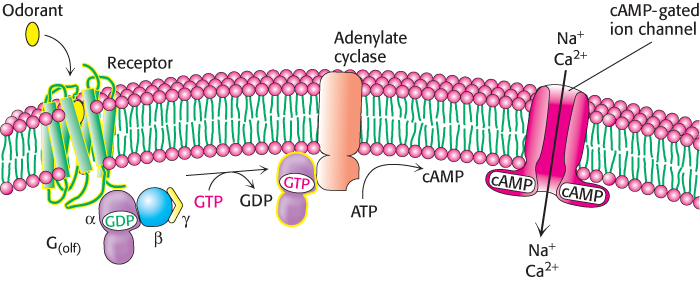
Figure 33.5: The olfactory signal-transduction cascade. The binding of odorant to the olfactory receptor activates a signaling pathway similar to those initiated in response to the binding of some hormones to their receptors. The final result is the opening of cAMP-gated ion channels and the initiation of an action potential.
Odorants are decoded by a combinatorial mechanism
An obvious challenge presented to an investigator by the large size of the OR family is to match each OR with the one or more odorant molecules to which it binds. Exciting progress has been made in this regard. Initially, an OR was matched with odorants by overexpressing a single, specific OR gene in rats. This OR responded to straight-chain aldehydes, most favorably to n-octanal and less strongly to n-heptanal and n-hexanal. More dramatic progress was made by taking advantage of our knowledge of the OR signal-transduction pathway and the power of the polymerase chain reaction (PCR). A section of nasal epithelium from a mouse was loaded with the calcium-sensitive dye Fura-2 (Section 14.1). The tissue was then treated with different odorants, one at a time, at a specific concentration. If the odorant had bound to an OR and activated it, that neuron could be detected under a microscope by the change in fluorescence caused by the influx of calcium that takes place as part of the signal-transduction process. To determine which OR was responsible for the response, cDNA was generated from mRNA that had been isolated from single identified neurons. The cDNA was then subjected to PCR with the use of primers that are effective in amplifying most or all OR genes. The sequence of the PCR product from each neuron was then determined and analyzed.
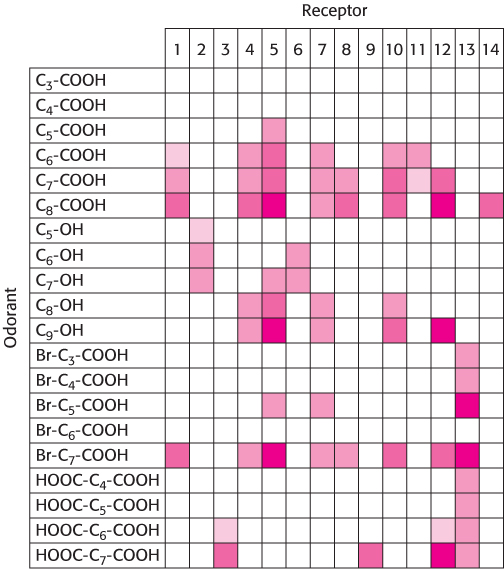
Figure 33.7: Patterns of olfactory-receptor activation. Fourteen different receptors were tested for responsiveness to the compounds shown in Figure 33.6. A colored box indicates that the receptor at the top responded to the compound at the left. Darker colors indicate that the receptor was activated at a lower concentration of odorant.
Using this approach, investigators analyzed the responses of neurons to a series of compounds having varying chain lengths and terminal functional groups (Figure 33.6). The results of these experiments appear surprising at first glance (Figure 33.7). Importantly, there is not a simple 1:1 correspondence between odorants and receptors. Almost every odorant activates a number of receptors (usually to different extents) and almost every receptor is activated by more than one odorant. Note, however, that each odorant activates a unique combination of receptors. In principle, this combinatorial mechanism allows even a small array of receptors to distinguish a vast number of odorants.
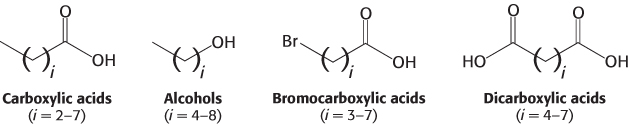
Figure 33.6: Four series of odorants tested for olfactory-receptor activation.
How is the information about which receptors have been activated transmitted to the brain? Recall that each neuron expresses only one OR and that the pattern of expression appears to be largely random. A substantial clue to the connections between receptors and the brain has been provided by the creation of mice that express a gene for an easily detectable colored marker in conjunction with a specific OR gene. Olfactory neurons that express the OR–marker-protein combination were traced to their destination in the brain, a structure called the olfactory bulb (Figure 33.8). The processes from neurons that express the same OR gene were found to connect to the same location in the olfactory bulb. Moreover, this pattern of neuronal connection was found to be identical in all mice examined. Thus, neurons that express specific ORs are linked to specific sites in the brain. This property creates a spatial map of odorant-responsive neuronal activity within the olfactory bulb.
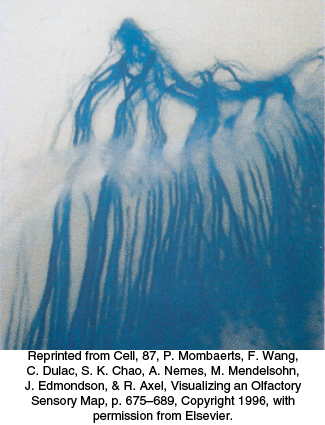
Figure 33.8: Converging olfactory neurons. This section of the nasal cavity is stained to reveal processes from sensory neurons expressing the same olfactory receptor. The processes converge to a single location in the olfactory bulb.
Can such a combinatorial mechanism truly distinguish many different odorants? An electronic “nose” that functions by the same principles provides compelling evidence that it can (Figure 33.9). The receptors for the electronic nose are polymers that bind a range of small molecules. Each polymer binds every odorant, but to varying degrees. Importantly, the electrical properties of these polymers change on odorant binding. A set of 32 of these polymer sensors, wired together so that the pattern of responses can be evaluated, is capable of distinguishing individual compounds such as n-pentane and n-hexane as well as complex mixtures such as the odors of fresh and spoiled fruit.

Figure 33.9: The Cyranose 320. The electronic nose may find uses in the food industry, animal husbandry, law enforcement, and medicine.