34.5 The Immune System Contributes to the Prevention and the Development of Human Diseases
The primary function of the immune system is to protect the host from invasion by foreign organisms. But how does the immune system prevent itself from mounting attacks against its host? In other words, how does the immune system distinguish between self and nonself? Clearly, proteins from the organism itself do not bear some special tag identifying them. Instead, selection processes early in the developmental pathways for immune cells kill or suppress those immune cells that react strongly with self-
T cells are subjected to positive and negative selection in the thymus
T cells derive their name from the location of their production—
Thymocytes produced in the bone marrow do not express the T-
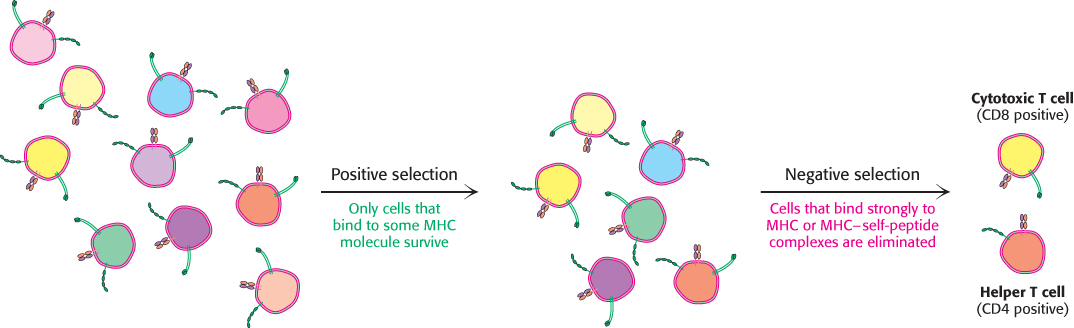
The cell population that survives positive selection is subjected to a second step, negative selection. Here, T cells that bind with high affinity to MHC complexes bound to self-
1005
Autoimmune diseases result from the generation of immune responses against self-antigens
 Although thymic selection is remarkably efficient in suppressing the immune response to self-
Although thymic selection is remarkably efficient in suppressing the immune response to self-
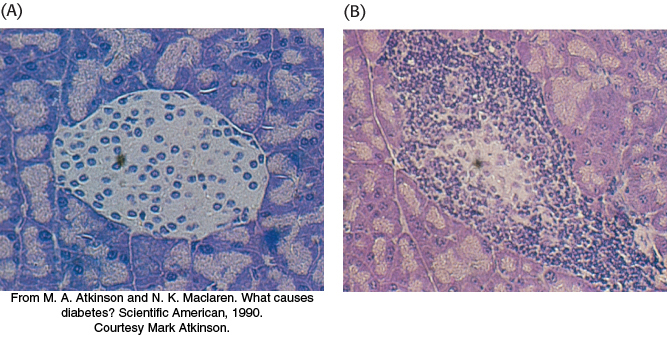
In many cases, the cause of the generation of self-
The immune system plays a role in cancer prevention
The development of immune responses against proteins encoded by our own genomes can be beneficial in some circumstances. Cancer cells have undergone significant changes that often result in the production of proteins that are not normally expressed. For example, the mutation of genes can generate proteins that do not correspond in amino acid sequence to any normal protein. Such proteins may be recognized as foreign, and an immune response will be generated specifically against the cancer cell. Alternatively, cancer cells often produce proteins that are expressed during embryonic development but are not expressed or are expressed at very low levels after birth. For example, a membrane glycoprotein called carcinoembryonic antigen (CEA) appears in the gastrointestinal cells of developing fetuses but is not normally expressed at significant levels after birth. More than 50% of patients with colorectal cancer have elevated serum levels of CEA. Immune cells recognizing epitopes from such proteins will not be subjected to negative selection and, hence, will be present in the adult immune repertoire. These cells may play a cancer surveillance role, killing cells that overexpress antigens such as CEA and preventing genetically damaged cells from developing into tumors.
1006
Vaccines are a powerful means to prevent and eradicate disease
The discovery and utilization of vaccines, biological preparations that stimulate immunity to a particular pathogen or disease, represents a significant milestone in modern medical history. The pioneering work of Edward Jenner and Louis Pasteur in the eighteenth and nineteenth centuries revealed that inoculation with inactivated forms of a pathogen could protect against subsequent infection with the active pathogen. Treatment of a person with a vaccine, or immunization, leads to the development of immunological memory, the ability of the immune system to respond more rapidly and effectively to pathogens that have been encountered previously. At the cellular level, memory arises because positively selected B cells and T cells that have a high affinity for their target persist within the organism as memory B cells and memory T cells.
In order for a vaccine to be effective, it must generate a sustained protective response appropriate for the targeted pathogen. For example, pathogens such as Clostridium tetani, the causative agent of tetanus, cause disease by the release of destructive proteins called toxins. Hence, an effective tetanus vaccine should generate a strong B-
Killed or inactivated vaccines contain pathogens that have been rendered harmless by treatment with chemicals or high heat.
Live attenuated vaccines contain live pathogens that have accumulated mutations so that they are no longer virulent to human cells. These vaccines are most commonly generated by repeated infection of the pathogen in cultured cells until its virulence is lost.
Subunit vaccines contain a purified protein component of the pathogen. Such proteins can be either isolated from infected material (such as blood from chronically infected patients) or generated by recombinant methods.
Toxoid vaccines are used against pathogens that employ an extracellular toxin to cause disease. These vaccines contain a form of the toxin that has been inactivated by treatment with chemicals or high heat.
The effectiveness of immunization efforts has made the global eradication of certain diseases possible. In 1966, 20 million cases of smallpox were reported worldwide. Just 14 years later, after coordinated vaccination efforts in regions of endemic disease, the World Health Assembly declared that smallpox had been eradicated. Nevertheless, despite the success of vaccines in the prevention of many devastating diseases, several pathogens have posed a significant challenge to vaccine development. For example, the development of an effective HIV vaccine has been complicated by the antigenic diversity of HIV strains. Because its mechanism for replication is prone to error, a population of HIV presents an ever-
1007