Chapter Introduction
16.1 Messenger RNA Capping and Polyadenylation
16.2 Pre-mRNA Splicing and Editing
16.3 RNA Transport and Degradation
16.4 Processing of Non-Protein-Coding RNAs
16.5 RNA Catalysis and the RNA World Hypothesis
MOMENT OF DISCOVERY

Melissa Jurica
One of the most exciting moments I recall was the first time I “saw” spliceosomes, using electron microscopy. I had been working for a long time to purify spliceosomes, the RNA-protein complexes that produce functional messenger RNAs by removing intervening sequences, or introns. No one knew what spliceosomes look like, and I hoped to use these samples to obtain structural information that would provide clues to the mRNA splicing mechanism. Unfortunately, my purified samples tended to clump together when spotted onto the copper grids used for sample analysis by electron microscopy, making the images I saw uninterpretable.
Then late one night, while working alone, I developed some micrographs of a new spliceosome sample that showed uniform individual particles that finally looked like macromolecules—the spliceosome! I was so excited and moved that I felt ready to cry—but there was nobody around to show my result to! I posted the wet negative of the micrograph on my advisor Nico Grigorieff’s door, and when he saw it the next morning we shared the joy of this breakthrough together.
The key to this result was a somewhat serendipitous discovery that the addition of EDTA, a magnesium ion–chelating agent, could suppress the tendency of the spliceosomes to aggregate. I think this is because SR proteins, which are associated with the spliceosome and tend to be sticky, require magnesium ions to bind to each other. Because we used EDTA to remove magnesium ions, the SR proteins lost the ability to associate tightly with the spliceosomes or each other. I still recall that overwhelming feeling of excitement mixed with happiness, and I use it as a well of motivation to this day, as my lab continues to push forward studies of the spliceosome.
—Melissa Jurica, on determining the first electron microscopic structures of spliceosomes
Our genome is not as large as we once imagined. Humans have only about 20,000 protein-coding genes, more than a fruit fly (14,000) and fewer than a rice plant (38,000). It turns out that much of the complexity in multicellular organisms is derived not from the number of genes but from the number of different proteins produced and the cell’s ability to regulate gene expression at the level of RNA. Some bacterial RNA molecules, and virtually all eukaryotic RNAs, are synthesized as biologically inactive precursors that must be chemically modified during or immediately following transcription. These precursors, called primary transcripts, are processed into mature, biologically functional molecules by specialized enzymes. RNA processing events are some of the most interesting molecular activities in RNA metabolism, because they govern how, when, and whether the RNA will be used in the cell.
Although many different kinds of RNA are processed, all processing events consist of the same sort of chemical reactions: cleavage and/or joining of RNA strands and, sometimes, modification of the nucleotides themselves. RNA processing is an active and engaging area of research, because these seemingly simple reactions lie at the heart of gene expression and gene regulation, and when they go awry, diseases can result. For example, errors in splicing can lead to cystic fibrosis and spinal muscular atrophy.
Studies of RNA processing mechanisms have also shed new light on evolution and the origin of life. Some of these processing reactions are catalyzed by RNAs, not by proteins, as was once believed. The discovery of catalytic RNAs and the ability of RNA to catalyze cleavage and ligation reactions in other RNAs has provided support for the theory that life began with an RNA molecule.
RNA metabolism is much more complex in eukaryotes than in bacteria (Figure 16-1). Most bacterial mRNAs are not processed before being translated into protein. Indeed, they are sometimes translated even while transcription is occurring. In contrast, eukaryotic mRNAs are chemically modified in the nucleus before being transported into the cytoplasm for translation on the ribosomes (located in the cytosol or on the endoplasmic reticulum; see Chapter 18). A protective cap, made from a modified guanosine, is added to the 5′ end of the primary transcript. And the 3′ end is modified by cleavage and the addition of a string of adenylate residues to create a 3′ poly(A) “tail.”
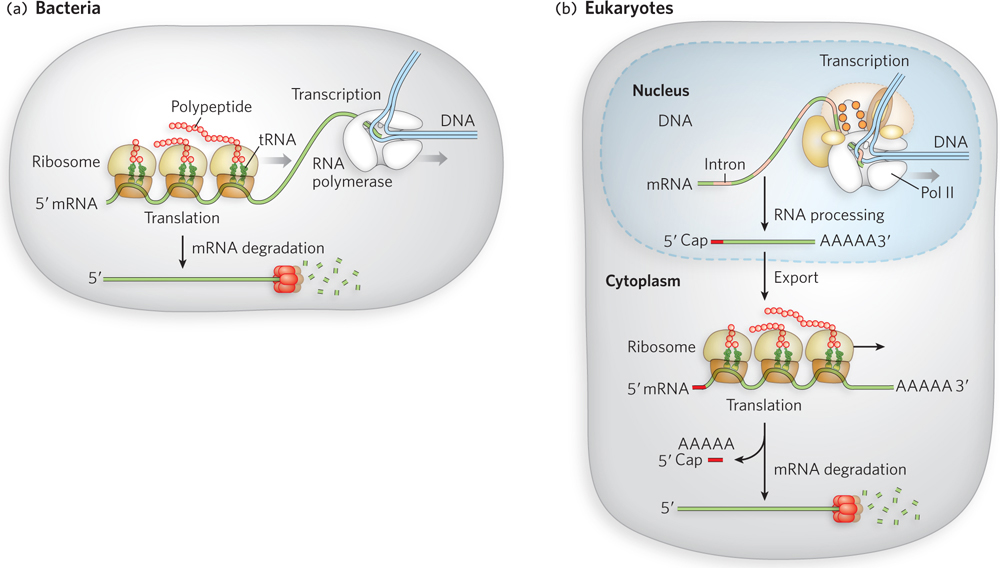
Figure 16-1: Messenger RNA processing in bacteria and eukaryotes. Eukaryotic mRNA undergoes several more steps than bacterial mRNA before translation. (a) Because bacterial cells lack a nucleus, transcription and translation are coupled. (b) In eukaryotic cells, transcription and pre-mRNA (precursor mRNA) processing occur in the nucleus, followed by export to the cytoplasm and subsequent translation. In both types of cells, the mRNA is eventually degraded.
Each primary transcript for a eukaryotic mRNA typically contains sequences encompassing just one gene, although the segments encoding the polypeptide may not be contiguous. Noncoding regions that interrupt the coding region of the transcript are known as introns, and the coding segments, as well as 5′ and 3′ untranslated regions (UTRs), are known as exons. In the process of splicing, the introns are removed from the primary transcript and the exons are covalently connected (ligated) to form a continuous sequence that specifies a functional polypeptide.
The enzymes that catalyze each of these three mRNA processing reactions—capping, polyadenylation, and splicing—do not operate independently. Instead, they seem to interact with one another and with the phosphorylated C-terminal domain (CTD) of RNA polymerase II (see Figure 15-23), such that each enzyme affects the function of the others. Processing of the transcript is also coupled to its transport from the nucleus to the cytoplasm. In this way, the cell keeps track of mRNAs that have been properly processed and can immediately put them to use for translation into protein. Once an mRNA transcript has reached the end of its utility in the cell, it is subject to a final type of RNA processing—degradation. Specific enzymes remove the 5′ cap and the 3′ poly(A) tail, and the rest of the RNA is broken down by cleavage of the phosphodiester backbone.
In this chapter, we examine how and when RNA molecules are processed and how these processes are coupled. We begin with mRNA processing, one of the earliest-discovered and most ubiquitous kinds of precursor-to-mature RNA transformation that occurs in cells, describing the mechanisms of capping, polyadenylation, and splicing, and a fourth type of RNA modification known as RNA editing. We then turn our attention to the transport of mRNA from the nucleus to the cytoplasm and eventual RNA degradation. We discuss the processing of non-protein-coding RNAs, including tRNA, rRNA, and tiny regulatory RNAs called microRNAs, and, finally, RNA catalysis and its implications for life’s origins.