Chapter Introduction
19: Regulating the Flow of Information
665
19.1 Regulation of Transcription Initiation
19.2 The Structural Basis of Transcriptional Regulation
19.3 Posttranscriptional Regulation of Gene Expression
MOMENT OF DISCOVERY

One of the most exciting moments in my career happened when I was working as a postdoctoral fellow with Greg Hannon at the Cold Spring Harbor Laboratories. We wondered whether microRNAs—
To test whether miR-
After many anxious hours of waiting, we got the results the next morning: all the tumor cells were green! This was incredibly exciting because it indicated that these tumors came from cells that overexpressed the miR-
—Lin He, on discovering that microRNA overexpression accelerates tumor development
666
All cells, whether single-
The relative amounts of each gene product required by a cell can also vary considerably. In an actively growing cell, for example, ribosomes are in high demand and can account for almost half of the cell’s dry weight, whereas only a few molecules of certain DNA repair proteins are required to do the necessary repair job. From an energy standpoint, having all gene products present in the highest possible amounts at all times would overburden the cell, considering the resources required to synthesize RNA and protein. Therefore, the expression of genes must be regulated so that their products are present in the right amount and only when they are needed.
Cells have evolved to respond to environmental changes and to adapt quickly to new growth conditions. This is how organisms can colonize a wide variety of biological habitats. Changing conditions range from variation in the availability and type of food sources to complex developmental regulatory programs in multicellular organisms, for which some gene products may be needed for a surprisingly brief time and in only a few cells.
Gene regulation is also important to the prevention of diseases, including cancer. In multicellular organisms, selective cell proliferation and destruction are critical for maintaining the proper levels of each cell type. The cell division cycle and programmed cell death pathways are exquisitely controlled by genes that promote or prevent these processes in response to cellular signals. When these regulatory genes are compromised, uncontrolled cell division can lead to the development of tumors. For example, p53 is a regulatory gene that plays a role in initiating programmed cell death. Loss-
Gene expression can be regulated at many different points in the synthesis of a functional RNA or protein. Transcription initiation is the most widely used regulatory point in both bacteria and eukaryotes, as this is the least costly way to control a gene. Initiation of transcription occurs at the very beginning of the synthetic pathway, before the investment in energy needed to make either RNA or protein. Nevertheless, mechanisms to regulate gene expression are found at virtually every point along the biosynthetic pathway. The points of regulation, shown in Figure 19-1, include (1) transcription initiation, (2) posttranscriptional processing (RNA processing), (3) RNA stability, (4) translation (protein synthesis), (5) protein modification, (6) protein transport, and (7) protein degradation.
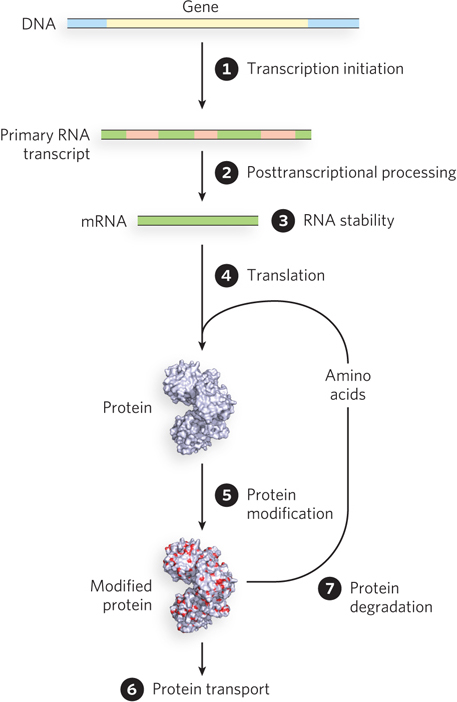
It is obviously important for cells to use their resources efficiently and not to waste energy synthesizing gene products that they do not need in a particular growth environment. But just as critical as efficiency is adaptability, and thus control: cells must be able to respond rapidly to changes in the need for various gene products. In other words, cells are control freaks! One could argue that control is ultimately more important than energy efficiency for the cell’s adaptation and survival. Although this point is often lost in discussions of gene regulation, it is central to biology. As you can see from Figure 19-1, and as we discuss in this and the following chapters, some regulatory mechanisms are directed at mRNA or even at the protein products of mRNA translation. Why do cells “waste” their efforts in this way? Such pathways provide a means of rapidly altering the levels of active proteins in response to the cell’s needs. Over the course of evolution, cells and organisms with such capability have won out over those that may have been more energy efficient but were less able to adapt to changing conditions. Thus, gene regulation involves a fine balance between efficiency and adaptability. New and surprising regulatory mechanisms continue to be discovered, and newly discovered posttranscriptional and translational regulatory processes are proving to be highly important, especially in eukaryotes.
667
In Chapters 15–18 we learned about the mechanics of transcription and translation, processes critical to the flow of biological information. Now we consider how these processes are regulated by the cell to conserve resources while effectively responding to changing environmental conditions and achieving optimum cell function. This chapter presents some general principles of gene regulation that are common to the mechanisms used by both bacteria and eukaryotes. We start by examining the protein-