2.3 The Brain
The brain has evolved from the brain stem structures that link the brain to the spinal cord all the way up to the cerebral cortex. As we go up the brain stem, processing gets more complex. In fact, it is the very top, the cerebral cortex, that differentiates our brains from those of all other animals. The cerebral cortex enables such complex processes as decision making, language, and perception. Even so, all of the structures below the cerebral cortex are essential for normal behavior and mental processing. This will become clearer as we discuss each structure’s role in this complex, interactive system we call the brain.
Going Up the Brain Stem
66
Between the spinal cord and the cortex, there are two sets of brain structures—
medulla A brain stem structure involved in many essential body functions, such as heartbeat, breathing, blood pressure, digestion, and swallowing.
pons A brain stem structure that serves as a bridge between the cerebellum and the rest of the brain and is involved in sleep and dreaming.
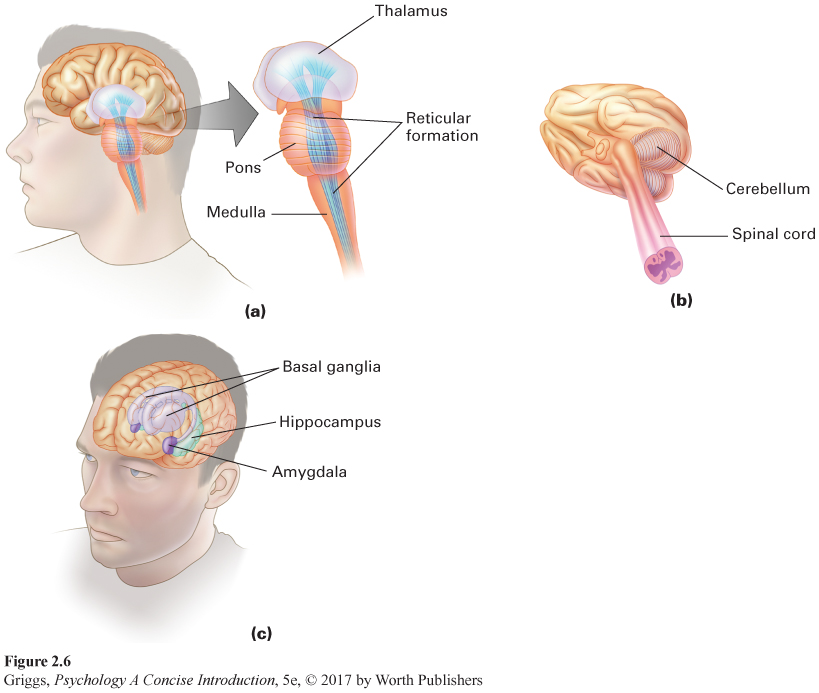
The central core. Figure 2.6 shows the central core brain structures. The brain stem spans from the spinal cord up to the thalamus. The first brain stem structure is the medulla, which links the spinal cord to the brain. The medulla is involved in regulating essential body functions such as heartbeat, breathing, blood pressure, digestion, and swallowing. This is why damage to the medulla can result in death. A drug overdose that suppresses proper functioning of the medulla can also lead to death. Just above the medulla, where the brain stem bulges, sits the pons. Along with the medulla, the pons serves as a passageway for neural signals to and from higher areas in the brain. The pons (Latin for “bridge”) functions as a bridge between the cerebellum and the rest of the brain and is involved in sleep and dreaming.
67
reticular formation A network of neurons running up the center of the brain stem that is responsible for our different levels of arousal and consciousness.
The reticular formation, a network of neurons running up the center of the brain stem and into the thalamus, is involved in controlling our different levels of arousal and awareness. This function was demonstrated by Moruzzi and Magoun (1949) in research with cats. When they stimulated a sleeping cat’s reticular formation, the cat awoke and entered a very alert state. When they severed the connections of the reticular formation with higher brain structures, however, the cat went into a coma from which it could never be awakened. The reticular formation also plays a role in attention by deciding which incoming sensory information enters our conscious awareness.
cerebellum A part of the brain involved in the coordination of our movements, sense of balance, and motor learning.
The cerebellum is involved in the coordination of our movements, our sense of balance, and motor learning. Cerebellum means “little brain” in Latin, and it looks like two minihemispheres attached to the rear of the brain stem. The cerebellum has roughly the same surface area (if unfolded) as a single cerebral hemisphere does (Bower & Parsons, 2007), but it is composed of smaller neurons so it takes up far less space. Only making up about 10% of the brain’s weight (Rilling & Insel, 1998), it has more neurons, an estimated 70 billion (Azevedo et al., 2009), than the rest of the brain combined. This is because the great majority of cerebellar neurons are granule neurons, which are very small and can be densely packed into a smaller space. The cerebellum coordinates all of our movements, such as walking, running, and dancing. Damage to the cerebellum will lead to very unsteady, unbalanced movement. Alcohol depresses the functioning of the cerebellum, leading to the uncoordinated movements typical of someone who is drunk. This is why some of the tests for being drunk involve coordinated movement. In addition, the cerebellum is the location of motor learning, such as how to ride a bicycle or to type. There is also some emerging evidence indicating that the cerebellum not only coordinates movement but may also play a role in integrating and coordinating sensory input and in mental functions such as planning (Bower & Parsons, 2007). Lastly, recovery from cerebellar damage, and even from removal of the cerebellum at a young age, is relatively good. Why this is so remains unanswered.
thalamus A part of the brain that serves as a relay station for incoming sensory information.
basal ganglia A part of the brain that is involved in the initiation and execution of movements.
The thalamus, located at the top of the brain stem, serves as a relay station for incoming sensory information. As such, it sends each type of sensory information (visual, auditory, taste, or touch) to the appropriate location in the cerebral cortex. The only type of sensory information that does not pass through the thalamus is olfactory (smell) information. Smell information goes directly from the receptors in our nose to the cortex. The basal ganglia are on the outer sides of the thalamus and are concerned mainly with the initiation and execution of physical movements. Like the cerebellum, the basal ganglia are affected by alcohol, and so make the movements required by tests for drunken driving difficult to execute. The basal ganglia are actually a group of various interacting brain regions. As we discussed earlier in this chapter, abnormally low dopamine activity in one region of the basal ganglia results in Parkinson’s disease. Another disease that involves difficulty in controlling movements is Huntington’s chorea, which stems from problems in another region of the ganglia in which there are GABA and acetylcholine deficits.
68
All of the various central core structures are summarized in Table 2.3 along with some of their major functions.
| Central Core Structure | Functions |
|---|---|
| Medulla | Involved in essential body functions, such as heartbeat, breathing, blood pressure, and swallowing |
| Pons | Serves as bridge between cerebellum and rest of brain and involved in sleep and dreaming |
| Reticular Formation | Responsible for our different levels of arousal and consciousness |
| Cerebellum | Involved in coordination of our movements, sense of balance, and motor learning |
| Thalamus | Serves as relay station for incoming sensory stimuli (except for olfactory sensory information) |
| Basal Ganglia | Involved in initiation and execution of movements |
limbic system A group of brain structures (hypothalamus, hippocampus, and amygdala) that play an important role in our survival, memory, and emotions.
hypothalamus A part of the brain that is involved in regulating basic drives such as eating, drinking, and having sex. It also directs the endocrine glandular system through its control of the pituitary gland and the autonomic nervous system to maintain the body’s internal environment.
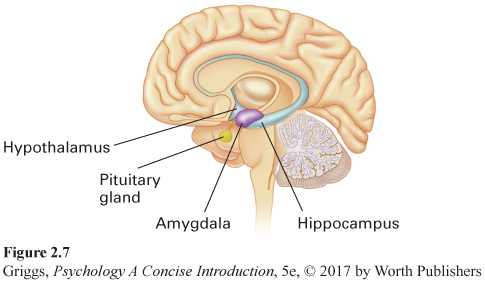
The limbic system. Surrounding the top border (or limbus in Latin) of the brain stem is the limbic system, which is made up of the hypothalamus, the amygdala, and the hippocampus. These limbic structures play an important role in our survival, memory, and emotions. Figure 2.7 shows the three parts of the limbic system. The hypothalamus is a very tiny structure, weighing about half an ounce, which is named after its location—
hippocampus A part of the brain involved in the formation of memories.
69
The hippocampus is involved in the formation of memories. Hippocampus may seem like a rather strange name, but like many parts of the brain, it was given a name that matched its visual appearance. The hippocampus looks somewhat like a seahorse, and hippocampus means “seahorse” in Greek. Recall the case study of H. M. that we discussed in Chapter 1. H. M. had his left and right hippocampal structures removed for medical reasons and as a result suffered severe memory deficits (Corkin, 1984). Subsequent research on H. M. and other people with amnesia has shown the hippocampus to be critical for the formation of certain types of new memories (Cohen & Eichenbaum, 1993), which we will discuss in Chapter 5.
The hippocampus has also been found to have the capacity to generate new neurons, a process called neurogenesis (Gage, 2003; Kempermann & Gage, 1999). The research group most responsible for demonstrating neurogenesis in humans was led by Fred Gage. Given that brain-
amygdala A part of the brain that is involved in emotions by influencing aggression, anger, and fear and by providing the emotional element of our memories and the interpretation of emotional expressions in others.
Located just in front of the hippocampal structures are the amygdala left and right structures. Amygdala means “almond” in Greek, and these structures look like almonds. The amygdala plays a major role in regulating our emotional experiences, especially fear, anger, and aggression. It is also responsible for generating quick emotional responses directly, without cortical involvement (LeDoux, 2000). The first evidence for the amygdala’s role in emotional behavior was done on wild, rather violent rhesus monkeys (Klüver & Bucy, 1939). The monkeys’ amygdalas were surgically removed. The surgery transformed the monkeys into calm, tame animals, clearly changing their emotional behavior. Other research has indicated that the amygdala also provides the emotional element in our memories and guides our interpretation of the emotional expressions of others (LeDoux, 1996).
70
All three structures in the limbic system are summarized in Table 2.4 along with some of their major functions.
| Limbic System Structure | Functions |
|---|---|
| Hypothalamus | Involved in regulating basic drives such as eating, drinking, and having sex; directing the endocrine glandular system; and maintaining the body’s internal environment |
| Hippocampus | Involved in the formation of memories and via neurogenesis may play a role in depression |
| Amygdala | Involved in emotions by influencing aggression, anger, and fear and by providing emotional element of our memories and the interpretation of emotional expressions in others |
Processing in the Cerebral Cortex
cerebral cortex The layers of interconnected cells covering the brain’s two hemispheres. This is the control and information-
corpus callosum The bridge of neurons that connects the two cerebral hemispheres.
All of the brain structures that we have discussed so far are important to our behavior and survival. The most important brain structure, however, is the cerebral cortex, the control and information-
The cerebral cortex is very crumpled in appearance with all sorts of bulges and gaps. This allows more cortical surface area to fit inside our rather small skull. If we were to unfold the cerebral cortex to check the amount of surface area, we would find the area to be about the size of four sheets of notebook paper or one newspaper page. Think about this. You couldn’t fit four sheets of paper into your pants pocket unless you crumpled them up. This is the same principle that applies to fitting the large surface area of the cerebral cortex into the small space within the skull. It is this large amount of surface area in the cerebral cortex that not only allows our complex cognitive processing but also differentiates our brains from those of all other animals. To see where different types of processing occur in the cerebral cortex, we need to learn the geography of the two hemispheres. This geography is rather simple in that the outer surface of each hemisphere is divided into four defined parts, called lobes, which we will discuss first.
71
frontal lobe The area in each cerebral hemisphere in front of the central fissure and above the lateral fissure. The motor cortex is in this lobe.
parietal lobe The area in each cerebral hemisphere in back of the central fissure and above the lateral fissure. The somatosensory cortex is in this lobe.
temporal lobe The area in each cerebral hemisphere located beneath the lateral fissure. The primary auditory cortex is in this lobe.
occipital lobe The area located in the lower back of each cerebral hemisphere. The primary visual cortex is in this lobe.
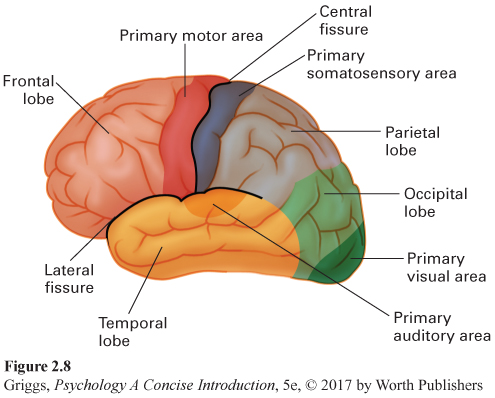
The four lobes. Figure 2.8 shows the four lobes in the left hemisphere. They are the same in the right hemisphere and are named after the specific bone area in the skull covering each of them. Two distinctive fissures (gaps) serve as boundary markers for three of the lobes. The central fissure (also called the fissure of Rolando) runs down the center of each hemisphere, and the lateral fissure (also called the Sylvian fissure) runs back along the side of each hemisphere. The four lobes are named for the four bones of the skull that overlie them. The frontal lobe is the area in front of the central fissure and above the lateral fissure, and the parietal lobe is the area located behind the central fissure and above the lateral fissure. The temporal lobe is located beneath the lateral fissure. The remaining lobe is the occipital lobe, which is located in the lower back of each hemisphere. Brynie (2009) explains how to find your occipital lobes. Run your hand up the back of your neck until you come to a bump and then place your palm on the bump. Your palm is squarely over the occipital lobes. The frontal lobes are the largest of the lobes. Their location is easy to remember because they are in the front of the hemispheres and directly behind the forehead. Now that we know these four areas, we can see what we have learned about what type of processing occurs in each of them. We begin with the well-
72
motor cortex The strip of cortex in each cerebral hemisphere in the frontal lobe directly in front of the central fissure, which allows us to move different parts of our body.
The motor cortex. The motor cortex, the frontal lobe strip of cortex directly in front of the central fissure in each hemisphere, allows us to move different parts of our body. The two motor cortex strips are related to the body by contralateral control—
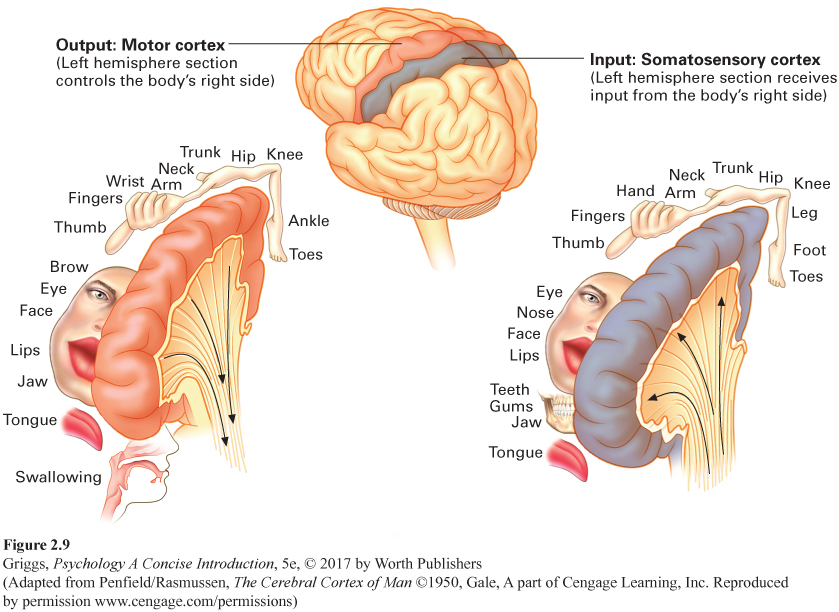
somatosensory cortex The strip of cortex in each cerebral hemisphere in the parietal lobe directly in back of the central fissure, which allows us to sense pressure, temperature, and pain in different parts of our body as well as the position of our body parts.
The somatosensory cortex. The somatosensory cortex, the parietal lobe strip of cortex directly behind the central fissure in each hemisphere, is where our body sensations of pressure, temperature, limb position, and pain are processed. Somato is Greek for body. Somatosensory, then, refers to the body senses. In addition to information about touch, the somatosensory cortex receives input about temperature and pain and information from the muscles and joints that allow us to monitor the positions of the various parts of the body. As in the motor cortex, there are contralateral relationships between the somatosensory strips and sides of the body. The somatosensory strip in the left hemisphere interprets the body sensory information for the right side of the body, and the strip in the right hemisphere interprets this information for the left side of the body. In addition, the amount of space within these strips is not allocated by the size of the body part. In the somatosensory cortex, it is allocated in accordance with the sensitivity of the body part—
73
You may be wondering how the homunculi for the motor and somatosensory strips were determined. They were the result of pre-
74
association cortex All of the cerebral cortex except those areas devoted to primary sensory processing or motor processing. This is where all the higher-
The visual cortex and the auditory cortex. There are two other important areas for processing sensory information in Figure 2.8 that we haven’t discussed yet. These are the primary processing areas for our two major senses, seeing and hearing. The visual cortex is located in the occipital lobes at the back of the hemispheres, and the auditory cortex is in the temporal lobes. Figure 2.8 shows where they are located in the left hemisphere. They have the same locations in the right hemisphere. The size of the primary visual cortex varies between individuals over a threefold range in surface area and volume (Dougherty et al., 2003). The reasons for this variability are not known. Interestingly, Schwartzkopf, Song, and Rees (2011) found a strong negative correlation between the magnitude of two context illusions and the surface area of the primary visual cortex. Thus, the smaller your primary visual cortex, the more dramatic some visual illusions appear. The primary visual cortex and the primary auditory cortex are only the locations of the initial processing of visual and auditory information. These primary areas pass the results of their analyses on to areas in the other lobes to complete the brain’s interpretation of the incoming visual or auditory information. These secondary cortical processing areas are part of what is termed the association cortex—all the areas of the cerebral cortex, except for those devoted to primary sensory or motor processing.
75
The four cerebral lobes and the type of sensory/motor processing that occurs in each lobe are summarized in Table 2.5.
| Cerebral Lobe | Type of Sensory/Motor Processing |
|---|---|
| Frontal | Includes motor cortex, which allows us to move the different parts of our body |
| Parietal | Includes somatosensory cortex where our body sensations of touch, temperature, limb position, and pain are processed |
| Temporal | Includes primary visual cortex where visual sensory information is initially processed |
| Occipital | Includes primary auditory cortex where auditory sensory information is initially processed |
Given the different locations of the visual cortex and the auditory cortex, you may think that seeing and hearing function entirely separately from each other. The reality, however, is that there is plenty of crosstalk between them. A good example of this interaction is the McGurk effect, which demonstrates that the brain integrates visual and auditory information when processing spoken language (McGurk & MacDonald, 1976). The McGurk effect arises out of conflicting auditory and visual information. A person’s lip movements influence what we hear. For example, if you listen to an audio track of a syllable (“ba”) spoken repeatedly while you watch a video clip of a person making the lip movements as if repeatedly saying another syllable (“ga”), you actually hear a third syllable “da,” the brain’s best guess after merging the information from the two senses. You will have a much better understanding of the McGurk effect if you experience it, and you can find several Web sites that allow you to do so just by doing an online search for the McGurk effect. The McGurk effect is a good example of how visual information can exert a strong influence on what we hear, but it is just one example of how our senses interact. There are many others, such as the smell of food influencing its taste. In interpreting the world, the brain blends the input from all of our senses.
It is important not to confuse sensory interaction in general and the McGurk effect, which all of us with normal sensory capabilities can experience, with synesthesia, a rare neurological condition in which otherwise normal people have cross-
76
The association cortex. Most of the cortex (about 70%) is association cortex. This is where all the higher-
Damage to the FFA has also been found to play a role in prosopagnosia (sometimes called face blindness), a condition that impacts a person’s ability to recognize faces including relatives, friends, and acquaintances or even oneself in a mirror (Greuter, 2007; Hadjikhani & de Gelder, 2002). Although a person with prosopagnosia cannot consciously recognize faces, some part of their brain does unconsciously distinguish familiar faces from unfamiliar ones because they show a higher skin conductance response to familiar faces (Tranel & Damasio, 1985). Sadly, prosopagnosia cannot be cured, but people with prosopagnosia learn other ways of recognizing people, such as memorizing people’s different speech patterns, their gaits, how they wear their hair, and so on. Surprisingly, prosopagnosia also occurs among people without brain damage and is highly heritable (Wilmer et al., 2010).
77
During the nineteenth century and most of the twentieth century, however, brain scans obviously weren’t available, so scientists had to learn about the association areas by other methods. One major method was to study the behavior of people who had suffered brain damage in accidents and strokes, or had brain surgery for medical reasons. Their behavioral and mental processing deficits were then related to the particular cortical areas where damage or surgery had occurred. A railroad worker’s tragic accident in 1848 gave us some hints as to what type of processing occurs in our frontal lobes (Macmillan, 2000). Phineas Gage was taking a break from tamping some blasting powder into a rock with a metal tamping iron. He was distracted and the tamping iron slipped, generating some sparks that caused the powder to explode. The explosion caused the metal tamping iron, which was 3 feet 7 inches long, 1¼ inches in diameter, and 13½ pounds in weight, to fly through his left cheek and head, exiting through his left frontal lobe (Ratiu, Talos, Haker, Lieberman, & Everett, 2004; Van Horn et al., 2012). Before Ratiu et al.’s study, most researchers believed that both frontal lobes were damaged, but Ratiu et al. showed that the cerebral injury was limited to the left frontal lobe. Their computerized tomography scan of Gage’s skull clearly shows this, and Van Horn et al. later verified the damage was localized to the left frontal lobe.
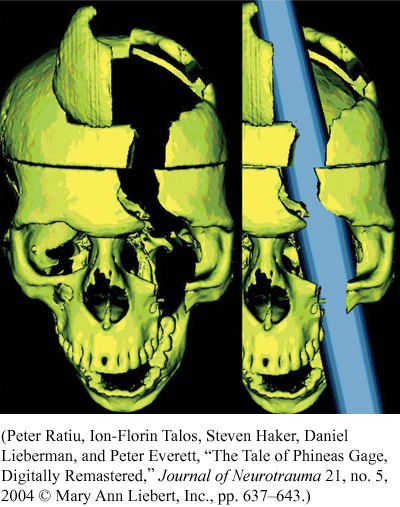
Gage survived, but his personality appeared to be greatly changed. According to his doctor and friends, Gage was “no longer Gage” (Macmillan, 2000). He became very irresponsible, impulsive, and disorderly; had great difficulty making decisions and reasoning; and cursed regularly. This led neuroscientists to hypothesize that the frontal lobes played a major role in such behaviors. Subsequent collaborative evidence from similar cases and from studies using other techniques such as brain scans confirmed that this is the case (Klein & Kihlstrom, 1998). The frontal lobes play a major role in our planning, decision making, judgment, impulse control, and personality, especially its emotional aspects. Actually, the cortical area destroyed in Gage’s left frontal lobe (the prefrontal cortex just behind the forehead and in front of the primary motor cortex) was essentially the left portion of the cortex that was disconnected from the rest of the brain in the infamous lobotomies performed on hundreds of patients in mental hospitals in the middle part of the twentieth century. The frontal lobes may also be partially responsible for the bad decisions and risky behavior of teenagers, because frontal lobe development is not complete until the late teens and possibly even the early 20s (Sabbagh, 2006).
78

What happened to Phineas Gage? While it has proved very difficult to fill in the exact details of Gage’s subsequent life, Fleischman (2002) provides a general account. Following his recovery, Phineas tried to return to his job as railroad foreman, but because of the “new” Gage’s vulgar, unreliable behavior, the railroad let him go. It appears that for a brief period of time he traveled around New England exhibiting himself along with the tamping iron, possibly ending up in P. T. Barnum’s American Museum sideshow in New York City. After this, because he got along very well with horses, he worked for a while in a livery stable in New Hampshire near his hometown. In 1852 he traveled, taking the tamping iron with him, to South America to care for horses and be a stagecoach driver in Chile. Because of health issues he returned in 1859 to California where his family had moved; and after recovering, he found work as a farmhand. In 1860 he began suffering epileptic seizures (likely due to slow changes in his brain tissue damaged in the railroad accident), and at that time, physicians could not control such seizures. These seizures killed him, 11½ years after the accident. The immediate cause of death was probably hypothermia created by the seizures (during which the body cannot control its internal temperature).
Macmillan and Lena (2010) provide a detailed argument that Gage seems to have made a surprisingly good psychosocial adaptation to his injury and that his mental changes were not as permanent as was once thought and that over time, he became a higher functioning, more socially adapted person. According to Macmillan and Lena, major factors in this rehabilitation were Gage’s highly structured employment and life over a period of nearly 8 years as a stagecoach driver in Chile and the social skills, responsibilities, and challenges entailed in this work. For example, driving a six-
79

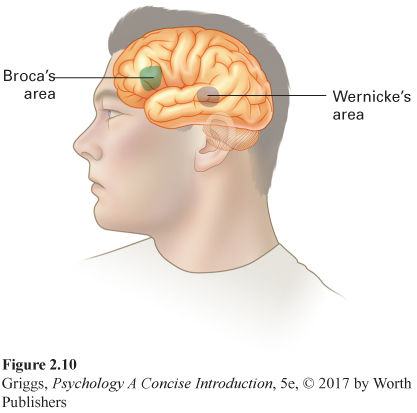
Broca’s area An area in the cerebral cortex responsible for fluent speech production. It is in the left frontal lobe of the majority of people, regardless of handedness.
Early brain researchers also analyzed the brains of deceased people, relating a person’s brain analysis to their behavior while living. Using this technique, two nineteenth-
80
Broca’s area is only in the left hemisphere in the majority of people, both right-
Wernicke’s area An area in the cerebral cortex responsible for comprehension of speech and text. It is in the left temporal lobe of the majority of people, regardless of handedness.
People with damage to Broca’s area do not have a problem understanding the speech of other people or reading silently, which means these skills must involve other areas. One of these areas is in the left temporal lobe and is named after its discoverer, Karl Wernicke, a German researcher. Wernicke’s area is responsible for the comprehension of speech and reading. Figure 2.10 shows exactly where this area is located. Damage to this area leads to Wernicke’s aphasia—
Like Broca’s area, Wernicke’s area is only in the left temporal lobe of the majority of people (regardless of handedness). Many people have the misconception that these speech and language centers normally reside in the hemisphere opposite to a person’s handedness. This is only the case for a very small number of right-
81

Broca and Wernicke examined the brains of deceased people who had speech and language dysfunctions in order to identify the speech and language centers in the brain. So perhaps this cortical localization technique could be used with highly intelligent people to identify the keys to their genius, the neural basis of intellect? What about the brain of acclaimed genius Albert Einstein? Was it studied after his death in 1955? The answer is a much qualified yes, because his brain was not studied in the careful, systematic manner that you would think it would be. The journey that Einstein’s brain took was indeed a bizarre odyssey (Abraham, 2001). With the exception of his brain and eyes, his body was cremated on the day of his death; but to this day no one seems to know where the ashes are (Lepore, 2001). The examining pathologist at Princeton Hospital where Einstein died, Dr. Thomas Harvey, removed the brain and took dozens of photographs of the whole and partially dissected brain before having it partitioned into 240 blocks of tissue, preparing a roadmap of the locations in the brain that yielded the dissected blocks, and sectioning and staining histological slides from the blocks.
Despite Harvey’s meticulous preservation of Einstein’s brain and the distribution of some brain sections to a few researchers, nothing had yet been published 23 years later when Harvey was interviewed in 1978 (Lepore, 2001). During this time period, Harvey kept most of the brain hidden away for his own personal research. He stored these remaining dissected and labeled brain pieces in glass jars of formaldehyde in a cider box in his office. Einstein’s brain usually accompanied him when he traveled. In fact, Dr. Harvey made a trip across America to visit Einstein’s granddaughter with some of the photographs and slides and some pieces of the brain floating in formaldehyde in a Tupperware bowl in a duffel bag in the trunk of a car (Paterniti, 2000).
Starting in the 1980s Harvey did dole out some more of the slides and photographs to researchers. This led to six peer-
82
Granted access to these materials, Falk et al. (2012) were able to describe the external gross neuroanatomy of Einstein’s entire cerebral cortex in comparison to 85 normal human brains. Many of the photographs show structures that were not visible in the photographs previously analyzed. Falk et al. found that Einstein’s brain was of unexceptional size but had very unusual morphology. The complexity and pattern of convolutions and folds in the prefrontal lobe, parietal lobe, visual cortex, and primary motor and somatosensory cortices gave these parts a much larger surface area than in normal brains. Falk et al. also speculated on the meaning of their findings. For example, the expanded prefrontal cortex may have contributed to the neurological substrates for some of Einstein’s remarkable abilities, and the unusual parietal lobes are consistent with the earlier hypothesis that they may have provided the neurological underpinnings for his visuospatial and mathematical abilities. But the long, strange journey for Einstein’s brain hasn’t ended. In 2012, digital preservations of some of the slides of Einstein’s brain became available to the general public as an iPad application. You have to wonder whether Einstein would have wanted images of his brain sold to nonscientists for $9.99.
Specializations of the Left and Right Hemispheres
Next, we will consider the question of whether there are any hemispheric specializations other than speech, language, and face perception. To try to answer this question, researchers have examined patients who have had their corpus callosum, the cortical bridge between the two hemispheres, severed as a medical treatment for severe cases of epilepsy. William Van Wagenen performed the first split-
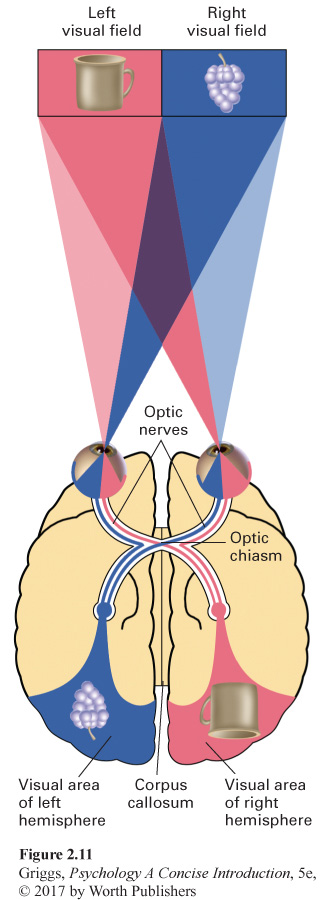
Studying the two hemispheres. To understand the experimental procedure for studying the two hemispheres, we first need to understand how the visual fields, the eyes, and the cerebral hemispheres are related. These relationships are shown in Figure 2.11. Our field of vision has two halves—
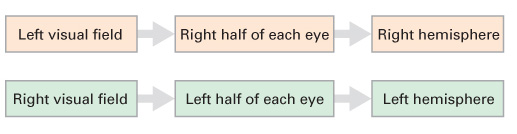
83
This relationship between the visual fields and the hemispheres is important because it allows information that is presented briefly in one visual field to go to only one hemisphere. With split-
Recall that the speech and language centers (Broca’s and Wernicke’s areas) are only in the left hemisphere of the vast majority of people. This would mean that a split-
84
Split-

What we know about the left and right hemispheres. The two most prominent researchers on split-

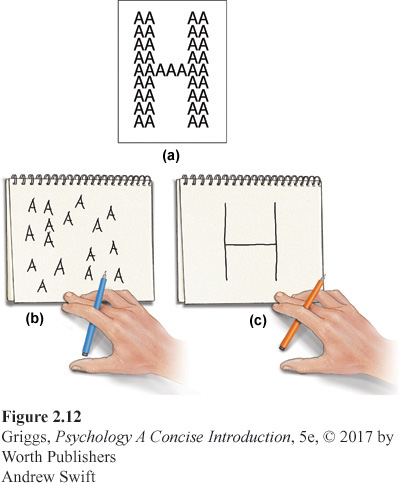
We know that the speech and language centers are specialties of the left hemisphere. This is why the left hemisphere is sometimes referred to as the verbal hemisphere. This does not mean, however, that the right hemisphere has no verbal ability. It can’t produce speech, but it can understand simple verbal material. In addition, the left hemisphere is better at mathematical skills and logic, while the right is better at spatial perception, solving spatial problems (e.g., block design), and drawing. The right hemisphere is also better at face recognition. Due to the asymmetry in the fusiform face area, recognition of faces is faster when faces are presented in the left-
85
Support for this general processing style difference between the hemispheres has been corroborated by research with brain-
86

This processing difference along with all of the other specialization differences that have been observed in split-
Consciousness and the Sleeping Brain
consciousness An individual’s subjective awareness of their inner thinking and feeling and their external environment.
There are many questions left for neuroscientists to answer about the brain and its many functions, but the most intriguing questions concern human consciousness—
Most brain (and bodily) functioning goes on without our conscious awareness. For example, you are presently recognizing the letters and words in this paragraph, but you are not consciously aware of the actual processing required to accomplish this recognition. You are only consciously aware of the results of the recognition process (the meaningful collection of words). Even if you wanted to, you couldn’t become consciously aware of this processing by the brain. Think about the billions of neurons communicating with one another within your brain and nervous system at this very moment. You cannot become aware of activity at this neuronal level of processing. Our consciousness is privy to only some of the results of this processing.
87
We can describe waking consciousness as our tuning in to both our internal and external worlds, but questions about exactly what underlies it (how it is generated) or why it evolved remain open for debate. Dehaene (2014) and Kaku (2014) provide some interesting theorizing about answers to these questions. Similarly, we are still pondering the question of why we experience sleep—
The five stages of sleep. The various stages of sleep have been determined by monitoring the electrical activity in the brain using an electroencephalograph (EEG). Small electrodes are attached to the scalp and connected to recording equipment that produces a real-
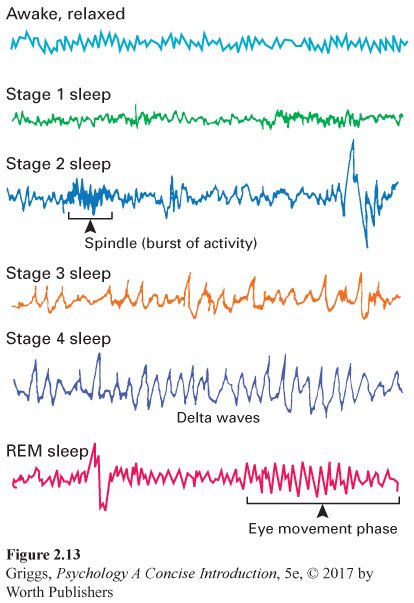
Next are the two stages composed of slow-
88
REM (rapid eye movement) sleep The stage of sleep that is characterized by rapid eye movements and brain wave patterns that resemble those for an awake state and in which most dreaming occurs. REM sleep is sometimes referred to as paradoxical sleep because the bodily muscles are immobilized but much of the brain is highly active.
Near the end of your first period of deep sleep (about an hour after falling asleep), you return through Stages 3 and 2 and enter REM (rapid eye movement) sleep, the most fascinating stage of sleep. REM sleep is characterized by very rapid waves somewhat like those of Stage 1 sleep. However, you are still sound asleep. REM sleep is sometimes referred to as paradoxical sleep because the muscles are relaxed but other body systems and the brain are active, following a waking pattern. Heart rate rises, breathing is more rapid, eyes move back and forth rapidly under closed eyelids, and one’s genitals become aroused. The brain is also highly active; both oxygen consumption and cerebral blood flow increase. Strangely, given all of this internal activity, the muscles are relaxed, leaving the body essentially paralyzed except for occasional muscle twitching.
REM sleep indicates the beginning of dreaming; it is the stage during which most dreaming occurs. How do we know this? If people are awakened during Stages 1 through 4, they only occasionally report dreaming; but if awakened during REM sleep, people usually recall dreams that they were having. This is as true for people who say they dream as for those who say they rarely do so, which means that we all dream whether we realize it or not. This first bout of REM sleep typically lasts about 10 minutes and completes the first sleep cycle of the night. The cycle of stages, in which we move out of and back into REM sleep, repeats itself about every 90 minutes throughout the night. Deep sleep (Stages 3 and 4) gets briefer and eventually disappears, as Stage 2 and REM sleep periods get longer with each sleep cycle. Over the course of the night, you have four or five REM sleep periods, amounting to about 20% to 25% of your sleep time (Figure 2.14).
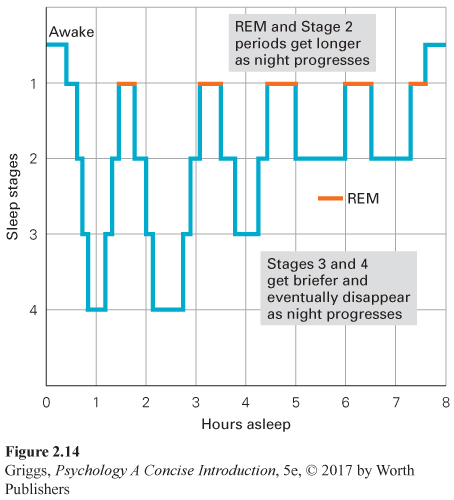
89
We spend on average about 2 hours a night dreaming, which works out to about 6 years of dreaming across our lifetime, given current life expectancies. We don’t remember many of our dreams, though. The dreams we do remember are usually from our last phase of REM sleep, right before we wake up. Many dreams are highly emotional and unpleasant. For example, people often dream of being attacked or pursued. This may be due to the fact that the frontal lobes (responsible for rational thought) are shut down during REM sleep, but the amygdala and the hippocampus, the limbic system structures responsible for emotion and memory, are active, creating the irrational imagery and emotional experiences of our dream world. Given the strong visual nature of dreams, what are the dreams of blind people like? Research indicates that people born blind or who go blind very early in life do not have visual dreams. Their dreams are just as emotional, but involve other sensations, especially sounds.
Why do we sleep and dream? We do not know why we sleep, but we know that we need sleep. You cannot cheat sleep; it will always overcome you. Inadequate sleep has a high cost. Sleep-
We clearly need sleep. How much, however, varies greatly with individuals. On the average, though, we spend about a third of our lives sleeping. Why? Four possible answers have been proposed, and the actual answer probably involves all four. First, sleep seems to serve a restorative function by allowing the brain and body time to rest and recuperate from the daily wear and tear of our existence. For example, recent research has shown that during sleep the brain cleans itself of toxic metabolic byproducts (Xie et al., 2013). This finding that sleep ensures metabolic homeostasis also has implications for many neurological disorders, many of which, such as Alzheimer’s disorder, are associated with sleep disturbances. These disturbances would prevent the brain from cleaning itself, and toxic byproducts would build up and cause brain damage. Second, gene activity during sleep ramps up the production of myelin, which as you have learned, protects our brain’s circuitry and facilitates neuronal transmission (Bellisi et al., 2013). Thus, sleep fortifies the brain over the long term by leading to the production of myelin, the brain’s insulating material. Third, sleep helps us process what we learn. It allows the brain the opportunity to consolidate our day’s experiences into our memory banks for later retrieval and use. Indeed, there is research evidence that memory is often strengthened and enhanced following a period of sleep (Stickgold & Ellenbogen, 2009). REM sleep is especially important in relation to memory. Disruption of REM sleep (by being constantly awakened during this stage) following learning impairs our memory for this learning (DuJardin, Guerrien, & LeConte, 1990). Disruption of other sleep stages does not similarly impair memory. Fourth, sleep may have evolved as an adaptive process. It served a protective function for our ancestors, increasing their survival by keeping them safe from predators during the night and from the dangers of wandering around cliffs in the dark. Those who stayed in their caves and slept had a better chance of surviving and passing on their genes.
90
So, sleep has clear advantages. But why do we dream? One answer is the critical role of REM sleep in learning—
In contrast to the activation-
91
Although sleep researchers do not agree about why we dream, they do agree that REM sleep is essential. This is clearly demonstrated by the REM sleep rebound effect—
Section Summary
In this section, we discussed many of the major structures in the brain. Between the spinal cord and the cerebral hemispheres, there are two sets of brain structures—
As we move up the brain stem to the cerebral hemispheres, the functioning gets more complex. It is the top cortical layer that gives us all of our complex abilities (such as language, perception, and decision making) and makes us “human.” Each cerebral hemisphere is divided into four lobes—
Research has shown that the frontal lobes play a major role in our planning, reasoning, impulse control, and personality—
92
Neuroscientists have much more to learn not only about the brain but also about consciousness, a person’s awareness of his inner thinking and feeling and his external environment. We have learned much about sleep, a natural altered state of consciousness, through the use of EEG recordings of the brain’s electrical activity while sleeping. Such recordings indicate the occurrence of five different stages of sleep, with the REM (rapid eye movement) sleep stage as the one during which most dreaming occurs. We can’t yet explain definitively why we sleep or why we dream, but both are clearly essential. Sleep-
3
Question 2.9
.
Explain where in the cerebral cortex (which hemisphere, lobe, area in the lobe, and part of that area) is active when you feel a slap on the left cheek of your face.
To feel a slap on the left cheek of your face, the area devoted to the cheek in the lower half of the somatosensory cortical strip in the parietal lobe in the right hemisphere would be activated. Refer to Figure 2.9 to see the exact location of the cheek in the somatosensory homunculus and thus the activation. Remember that the activation would be at this location in the right hemisphere because your left cheek was slapped.
Question 2.10
.
Explain which areas in the cerebral cortex are involved and how they are involved when you shadow (repeat aloud) someone’s speech.
When you shadow someone’s speech, the auditory sensory input from the person’s speech goes first to the primary auditory cortex in the temporal lobes, next to Wernicke’s area to be understood, and then to Broca’s area to produce the appropriate motor programs, which are then sent to the motor cortex, which then executes the programs to activate your speech muscles to produce the speech.
Question 2.11
.
Explain what types of behavioral deficits would be observed in a person who suffered damage to her cerebellum.
A person who suffered damage to her cerebellum would have problems in motor coordination and balance. Her movements would be jerky and uncoordinated, and she would have great difficulty maintaining her balance.
Question 2.12
.
Explain how you could demonstrate that a split-
You would ask him to identify the object by the sense of touch with his left hand. He could identify it by matching the tactile input (the information gathered from touching the object) with his knowledge of what had been presented, both in the right hemisphere.
Question 2.13
.

This 400-
If Vertumnus were flashed briefly in a split-
93
Question 2.14
.
Explain why REM sleep is sometimes referred to as paradoxical sleep.
REM sleep is sometimes referred to as paradoxical sleep because your bodily muscles are relaxed and immobilized, but many parts of your brain are active with both oxygen consumption and cerebral blood flow increasing. The brain appears to be awake, but you are behaviorally asleep.