4.3 Biological and Cognitive Aspects of Learning
In our discussion of the classical conditioning of emotional responses, we mentioned biological predispositions to learning certain fears. Humans seem prepared to learn fears of animals or heights much easier than fears of toy blocks or curtains. Animals (snakes or spiders) are more dangerous to us than toy blocks, so such predispositions make evolutionary sense (Seligman, 1971). Are there any other predispositions that affect classical conditioning and are there any biological constraints on operant conditioning? In addition to addressing these questions about the biological aspects of learning, we will also discuss cognitive research on latent learning and learning through modeling that questions whether reinforcement is necessary for learning. Let’s start with the biological research.
Biological Preparedness in Learning
Do you have any taste aversions? Have you stopped eating a certain food or drinking a certain liquid because you once got miserably sick after doing so? If you have a really strong aversion, you may feel ill even when you think about that food or drink. If the illness involved a specific food from a particular restaurant, you may have even generalized the aversion to the restaurant. Just as we are biologically prepared to learn certain fears more than others (Öhman & Mineka, 2001), we are also prepared to learn taste aversions. Our preparedness to learn to fear objects and situations dangerous to us (animals and heights) and to avoid foods and drinks that make us sick has adaptive significance. Such learning enhances our chances of survival. It makes biological sense then that we should be predisposed to such learning. To see how psychologists have studied such predispositions, let’s take a closer look at some of the early research on taste aversion.
185
Taste aversion. John Garcia and his colleagues conducted some of the most important early research on taste aversion in the 1960s. Their research challenged the prevailing behaviorist argument that an animal’s capacity for conditioning is not limited by its biology (Garcia, 2003). Garcia benefited from an accidental discovery while studying the effects of radiation on rats (Garcia, Kimeldorf, Hunt, & Davies, 1956). The rats would be moved from their home cages to experimental chambers for the radiation experiments. The radiation made the rats nauseated, and they would get very sick later back in their home cages. The rats, however, would still go back into the experimental chambers where they had been radiated, but they would no longer drink the water in these chambers. Why? The water bottles in the chambers were made of a different substance than those in the home cages—
So how did Garcia and his colleagues use these taste aversion results for rats to demonstrate biological preparedness in learning? Garcia and Koelling (1966) showed that the rats would not learn such aversions for just any pairing of cue and consequences. Those that seemed to make more biological sense (different-

186
This doesn’t mean that other animals might not be predisposed to learn auditory or visual aversions. For example, many birds, such as quail, seem to learn visual aversions rather easily. A clever study demonstrated this difference among animals in types of learning predispositions. In this study, both quail and rats drank dark blue, sour-
instinctual drift The tendency for an animal to drift back from a learned operant response to an innate, instinctual response to an object.
Instinctual drift. Keller and Marian Breland, two of Skinner’s former students, discovered an important biological preparedness effect on operant conditioning (Breland & Breland, 1961). The Brelands, who became animal trainers, employed operant conditioning to train thousands of animals to do all sorts of tricks. In doing this training, they discovered what has become known as instinctual drift—the tendency of an animal to drift back from a learned operant response to an object to an innate, instinctual response. For example, the Brelands used food reinforcement to train some animals to pick up oversized coins and put them in a bank. The Brelands did this with both pigs and raccoons. However, they observed that once the coins became associated with the food reinforcement, both types of animal drifted back to instinctual responses that were part of their respective food-

187
The important point of these findings is that biologically instinctual responses sometimes limit or hinder our ability to condition other, less natural responses. The Brelands’ work demonstrates a biological preparedness effect upon operant conditioning. Biological predispositions show that animals will learn certain associations (those consistent with their natural behavior) more easily than others (those less consistent with their natural behavior). Also note that this “misbehavior” of the pigs and the raccoons (their instinctual responses) continued without reinforcement from the trainers. In fact, it prevented the animals from getting reinforcement. This aspect of the Brelands’ work relates to the more general question of whether we can learn without reinforcement, which we will discuss in the next section.
Latent Learning and Observational Learning
Cognitively oriented learning researchers are interested in the mental processes involved in learning. These researchers have examined the question of whether we can learn without reinforcement in their studies of latent learning and observational learning. We’ll consider some of the classic research on these two types of learning.

latent learning Learning that occurs but is not demonstrated until there is incentive to do so.
Latent learning. Think about studying for an exam in one of your courses. What you have learned is not openly demonstrated until you are tested on it by the exam. You learn, but you do not demonstrate the learning until reinforcement for demonstrating it (a good grade on the exam) is available. This is an example of what psychologists call latent learning, learning that occurs but is not demonstrated until there is incentive to do so. This is what Edward Tolman was examining in his pioneering latent-
In this research, food-
188
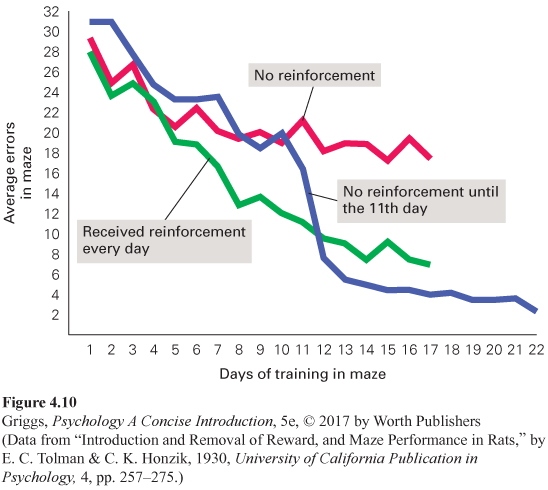

189
How would such a map be created in the rat’s brain and how would it allow the rat to navigate its way through a complex environment, such as a maze? A few decades elapsed after Tolman’s research before these questions were answered. The answer involves the discovery of two types of neurons in the brain, place cells and grid cells. Physiological psychologist John O’Keefe identified the place cells located in the hippocampus that function to build an inner map of the environment (O’Keefe, 1976; O’Keefe & Dostrovsky, 1971), and May-
observational learning (modeling) Learning by observing others and imitating their behavior.

Observational learning. Much learning by humans is more cognitive in nature and does not involve conditioning through direct experience. Observational learning, sometimes called modeling, is a good example. By watching a model, we learn through vicarious reinforcement and punishment without being conditioned directly. We learn to anticipate a behavior’s consequences in situations like those that we have observed. Observational learning (modeling)—learning by observing others and imitating their behavior—
Albert Bandura’s famous experiments on learning through modeling involved a Bobo doll, a large inflated clown doll, and kindergarten-
190

But what would happen if a child were exposed to a model that acted gently toward the Bobo doll? In another experiment, Bandura had children observe either an aggressive model, a gentle model, or no model. When allowed to play in a toy room that included a Bobo doll, what happened? The children exposed to the aggressive model acted more aggressively toward the doll than the children with no model, and the children with the gentle model acted more gently toward the doll than the children with no model. In general, the children’s behavior with the doll was guided by their model’s behavior. But the models in these experiments were not reinforced or punished for their behavior. Would this make a difference?
Bandura examined this question in another experiment. The young children watched a film of an adult acting aggressively toward the Bobo doll, but there were three different versions of the film. In one version, the adult’s behavior was reinforced; in another, the behavior was punished; and in the third, there were no consequences. Each child’s interactions with the Bobo doll varied depending upon which film the child had seen. The children who had watched the adult get reinforced for aggressive behavior acted more aggressively toward the Bobo doll than those who had seen the model act with no consequences. In addition, the children who had watched the adult get punished were less likely to act aggressively toward the doll than the children who had not been exposed to any consequences for acting aggressively toward the doll. The children’s behavior was affected by the consequences witnessed in the film. Then Bandura cleverly asked the children if they could imitate the behavior in the film for reinforcement (snacks). Essentially, all of the children could do so. This is an important point. It means that the children all learned the behavior through observation regardless of whether the behavior was reinforced, punished, or neither.
191
According to Lansford (2012), researchers subsequently questioned the generalizability of Bandura’s findings both with respect to the context (the studies were conducted in a laboratory setting) and the temporal proximity between the children’s observation of the adult model’s aggression and their own aggression (there was a close temporal proximity in the studies). Would children imitate aggression in other settings and after lengthier delays? It turns out that they do. More recent studies have found that Bandura’s finding of children imitating aggression does hold in a variety of nonlaboratory settings and after lengthy delays between observing the aggression and then acting aggressively (Bushman & Huesmann, 2010; Guerra, Huesmann, & Spindler, 2003; Slater, Henry, Swaim, & Anderson, 2003).
Much of the research on observational learning since Bandura’s pioneering studies has focused on the question of whether exposure to violence in media leads people to behave more aggressively. There is clearly an abundance of violence on television and in other media. It has been estimated that the average child has viewed 8,000 murders and 100,000 other acts of violence on television alone by the time he finishes elementary school (Huston et al., 1992), and more recent research indicates that the amount of violence on television and in other media has increased since Huston et al.’s study. For example, the Parents Television Council (2007) found that violence in prime-
Literally hundreds of studies have addressed this question, and as you would expect, the findings are both complicated and controversial. Leading scientists reviewing this literature, however, came to the following general conclusion: “Research on violent television and films, video games, and music reveals unequivocal evidence that media violence increases the likelihood of aggressive and violent behavior in both immediate and long-
192
The empirical evidence linking exposure to media violence to increased risk of aggressive behavior seems clear-
mirror neurons Neurons that fire both when performing an action and when observing another person perform that same action.
Recent research has also identified neurons that provide a possible neural basis for observational learning. These mirror neurons are neurons that fire both when performing an action and when observing another person perform that same action. When you observe someone engaging in an action, similar mirror neurons are firing in both your brain and in the other person’s brain. Thus, these neurons in your brain are “mirroring” the behavior of the person you are observing. Mirror neurons were first discovered in macaque monkeys via electrode recording by Giacomo Rizzolatti and his colleagues at the University of Parma in the mid-
193
Because both human and nonhuman primates learn much through observation and imitation, mirror neuron systems, if they function as their proponents claim they do, would provide a way through which observation could be translated into action (Cattaneo & Rizzolatti, 2009; Iacoboni, 2005, 2009b). Indeed, some studies claim that mirror neurons have proved useful in the rehabilitation of motor deficits in people who have had strokes (Ertelt et al., 2007). For example, patients who watch videos of people demonstrating various arm and hand movements (action observation) improved faster than those who did not watch the videos (Binkofski & Buccino, 2007). In addition to providing a neural basis for observational learning, some researchers have speculated that mirror neuron systems may also play a major role in both empathy (the ability to feel and understand the feelings of others) and understanding the intentions of others (Iacoboni, 2009a). In addition, mirror neurons have been proposed to be the basic neural mechanism from which language evolved (Rizzolatti & Arbib, 1998; Rizzolatti & Craighero, 2004). Some other researchers even think that there may be a link between mirror neuron deficiency and social disorders, namely autism, in which individuals have difficulty in social interactions (Dapretto et al., 2006). All of these claims for mirror neurons, especially the more speculative ones, such as the links to autism and language, are very controversial (Hickok, 2014; Jarrett, 2015). For example, a review of the results from 25 relevant studies concluded that there was little evidence for a global dysfunction of the mirror neuron system in autism (Hamilton, 2013). It is clear that most people with autism have no problem understanding other people’s actions and have normal imitation abilities and thus do not have dysfunctional mirror neuron systems (Gallese, Gernsbacher, Heyes, Hickok, & Iacoboni, 2011, p. 290). In sum, researchers not only disagree about the presence of mirror neuron systems in humans but also, if they do exist, about what exactly they do (Jarrett, 2015).
Lastly, observational learning, like latent learning, emphasizes the role of cognitive processes in learning. As demonstrated by our examples, just as Tolman’s rats seemed to have a cognitive map of the maze, Bandura’s children seemed to have a cognitive model of the actions of the adult models and their consequences. Cognitive psychologists have studied the development of such mental representations and their storage in memory and subsequent retrieval from memory in their attempts to understand how the human memory system works. We will take a detailed look at the human memory system in Chapter 5.
194
Section Summary
In this section, we learned about some of the effects of biological preparedness on learning and about latent learning and observational learning in which direct reinforcement is not necessary. Research on taste aversion indicated that rats could easily learn aversion to different-
Research has shown that reinforcement is not necessary for learning to occur. Tolman’s research with rats running mazes showed that without reinforcement, rats could learn a cognitive map of the maze that they could then use very efficiently when reinforcement became available at the end of the maze. This was an instance of latent learning, learning that is not demonstrated until there is an incentive to do so. Albert Bandura’s pioneering research on observational learning showed that much of human learning also doesn’t involve direct experience. The children in his study all learned a model’s behavior through observation, regardless of whether the behavior was reinforced, punished, or neither. Research in observational learning since Bandura’s studies has extended his findings by linking exposure to media violence to an increased likelihood of aggression in viewers. Other recent research by neuroscientists has led to the discovery of mirror neuron systems that may provide a possible neural basis for observational learning. However, their existence and functions in humans are controversial and have not been established by current research findings.
3
Question 4.10
.
Explain why the ease of learning taste aversions is biologically adaptive for humans.
Learning taste aversions quickly and easily is adaptive because it increases our chances of survival. If we eat or drink something that makes us terribly sick, it is adaptive to no longer ingest that food or drink because we might die. We have a greater probability of surviving if we learn such aversions easily.
Question 4.11
.
Given Garcia and Koelling’s (1966) findings (discussed in this section) for rats’ pairing the cues of sweet-
The rats easily learned (stopped drinking the water) when the normal-
Question 4.12
.
Explain why it would be easier to operantly condition a behavior that is “natural” for an animal than one that isn’t natural.
It would be easier to operantly condition a “natural” response because it would lower the probability that instinctual drift will interfere with the conditioning. Because an animal would already be making its natural response to the object, there would be no other response to drift back to. In addition, the natural response to the object would be easier to shape because it would be given sooner and more frequently than an unnatural response.
Question 4.13
.
Explain the relationship between latent learning and reinforcement.
Latent learning occurs without direct reinforcement, but such learning is not demonstrated until reinforcement is made available for the learned behavior.
Question 4.14
.
Explain how reinforcing and punishing models influenced observers in Bandura’s research.
In Bandura’s work, reinforcing the model increased the probability that the observed behavior would be displayed, and punishing the model decreased the probability that the observed behavior would be displayed. But Bandura demonstrated in both cases that the behavior was learned. The reinforcement or punishment only affected whether it was displayed.