Concept 5.5: The Membrane Plays a Key Role in a Cell’s Response to Environmental Signals
A hallmark of living cells is their ability to process information from their environments. We can think of this information in terms of cell signaling. In this context, the signal may be a physical stimulus such as light or heat, or a chemical such as a hormone. A chemical signal may also be referred to as a ligand: a molecule that binds to a receptor (see Concept 5.4). The mere presence of a signal, however, does not mean a particular cell will respond to it. In order to respond, the cell must have a specific receptor that can detect the signal. Once the signal activates its receptor, it sets off a signal transduction pathway, a sequence of molecular events and chemical reactions within a cell that lead to the cell’s response to the signal. This ability of cells to sense and respond to signals in the environment is key to the maintenance of stable intracellular conditions, a theme that recurs throughout this book.
Cells are exposed to many signals and may have different responses
Inside a large multicellular animal, chemical signals made by the body itself reach a target cell by local diffusion or by circulation within the blood. These signals are usually in tiny concentrations (as low as 10−10 M) and differ in their sources and mode of delivery (FIGURE 5.10):
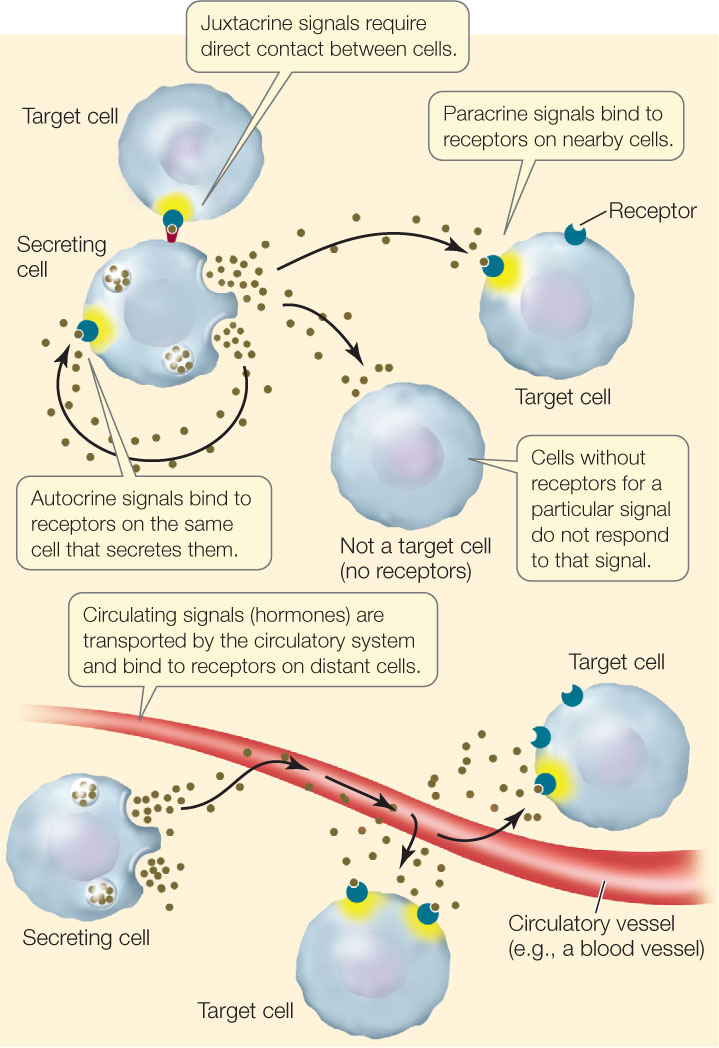
- Autocrine signals affect the same cell that releases them. For example, many tumor cells reproduce uncontrollably because they self-stimulate cell division by making their own division signals.
- Paracrine signals diffuse to and affect nearby cells. An example is a neurotransmitter made by a nerve cell that diffuses to an adjacent cell and stimulates it.
- Juxtacrine signaling requires direct contact between the signaling and the responding cell, and usually involves interaction between signaling molecules bound to the surfaces of the two cells.
- Signals that travel through the circulatory systems of animals or the vascular systems of plants to reach receptors on distant cells are generally called hormones.
Chemical signals do not always come from within the multicellular organism—some come from the external environment. For example, specific molecules produced by pathogenic (disease-causing) organisms trigger signal transduction pathways in plants, leading to defense responses.
For the information from a signal to be transmitted to a cell, the target cell must be able to sense the signal and respond to it. In a multicellular animal, all the cells may receive chemical signals that are circulated in the blood, but most body cells are not capable of responding to every signal. Only the cells with the necessary receptors can respond.
96
Typically, a signal transduction pathway involves a signal, a receptor, and a response (FIGURE 5.11). These pathways vary in their details, but they commonly include allosteric regulation. Recall that allosteric regulation involves an alteration in the three-dimensional shape of a protein as a result of the binding of another molecule at a site other than the protein’s active site (see Figure 3.20). You saw an example of allosteric regulation earlier in this chapter when we considered a ligand-gated channel, which opens (changes shape) after binding to another molecule (see Figure 5.4).
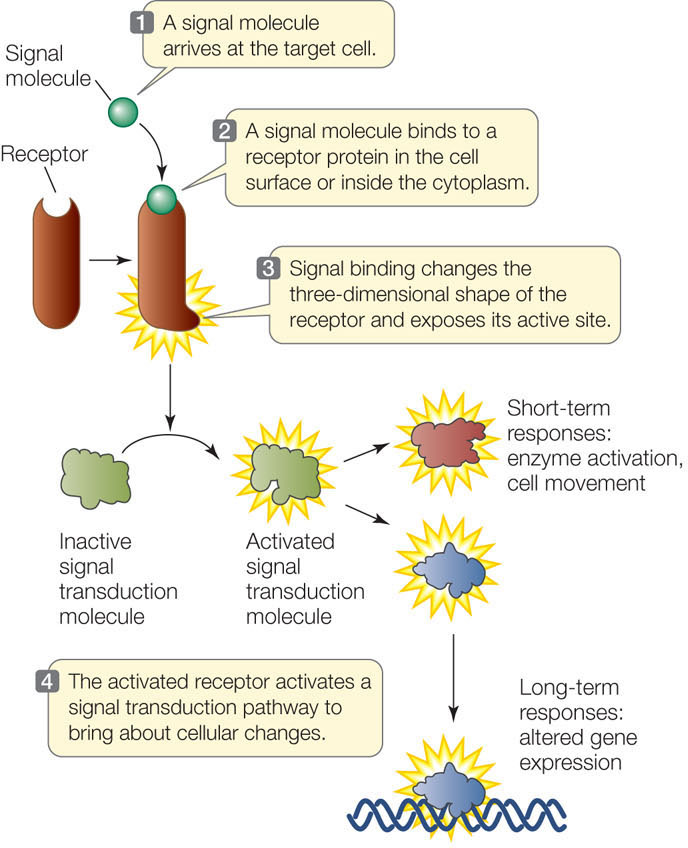
A signal transduction pathway may end in a response that is short-term, such as the activation of an enzyme, or long-term, such as an alteration in gene expression.
LINK
Gene expression—the transcription of specific DNA sequences and the translation of these sequences into proteins—is described in Chapter 10
Receptors can be classified by location and function
Chemical signals (ligands) are quite variable, but they can be divided into two groups based on whether or not they can diffuse through membranes. Physical signals such as light and sound also vary in their ability to penetrate particular cells and tissues. Accordingly, we can classify a receptor by its location in the cell, which largely depends on the nature of its ligand:
- Intracellular receptors are located inside the cell. Small or nonpolar ligands can diffuse across the phospholipid bicells. Acetylcholine layer of the cell membrane and enter the cell. Estrogen, for example, is a lipid-soluble steroid hormone that can easily diffuse across the cell membrane; it binds to a receptor inside the cell.
- Membrane receptors are located on the cell surface. Large or polar ligands cannot cross the lipid bilayer. Insulin, for example, is a protein hormone that cannot diffuse through the cell membrane. Instead, it binds to a transmembrane receptor with an extracellular ligand-binding domain.
Many receptors are associated with the cell membrane
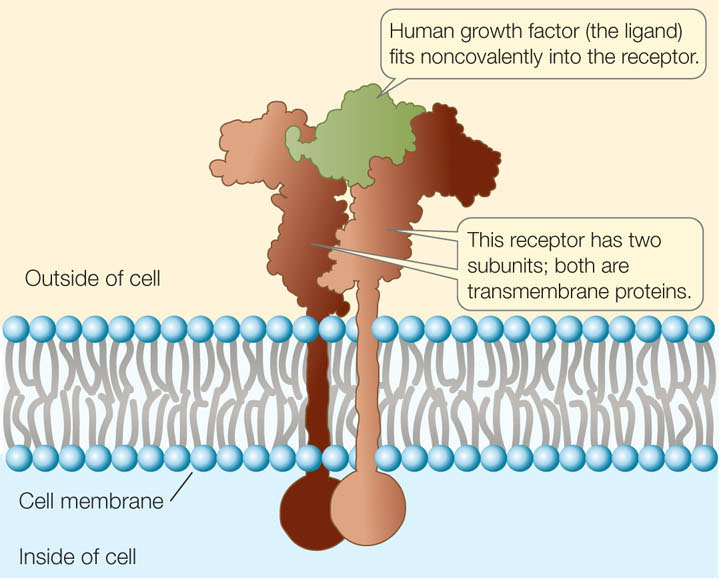
Membrane receptors are found on the surfaces of cells, and they respond to signals from outside the cell. If the signal is a chemical ligand, it fits into a three-dimensional site on its corresponding receptor protein (FIGURE 5.12). In many cases the receptor has a catalytic domain and functions as an enzyme, with its active site on the cytoplasmic side of the membrane. The ligand acts as an allosteric regulator, exposing the active site of the catalytic domain. The ligand does not contribute further to the cellular response; its role is purely to “knock on the door.” (This is in sharp contrast to the enzyme–substrate interactions we described in Concept 3.3. The whole purpose of those interactions is to change substrates into useful products.)
97
Ligands (L) bind to their receptors (R) noncovalently and reversibly, according to chemistry’s law of mass action:
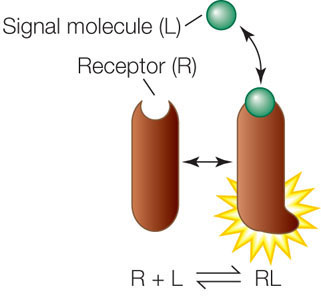
Reversibility is important because if the ligand were never released, the receptor would be continuously stimulated. In most cases, the cell needs to stop responding to a signal after the appropriate response has occurred. For example, if a signal transduction pathway results in the production of a particular protein, the cell needs to stop producing the protein when enough of it has been made. Nevertheless, for most ligand–receptor complexes, the equilibrium point is far to the right in the above reaction—that is, binding is favored, even at low ligand concentrations.
As noted above, receptors are removed from the cell membrane by endocytosis and either degraded inside the cell or recycled back to the membrane.
An inhibitor (or antagonist) can also bind to a receptor protein, preventing the binding of the normal ligand. This is analogous to the competitive inhibition of enzymes (see Concept 3.4). There are both natural and artificial antagonists of receptor binding. For example, many substances that alter human behavior (such as caffeine; see the opening story) bind to specific receptors in the brain and prevent the binding of the receptors’ specific ligands.
In complex eukaryotes such as mammals and higher plants, there are three well-studied categories of cell membrane receptors, which are grouped according to their activities: ion channels, protein kinase receptors, and G protein–linked receptors. Because you will see these receptors several times later in this book, we describe them in some detail here.
Ion Channel Receptors
As described in Concept 5.2, the cell membranes of many cells contain ligand-gated channels for ions such as Na+, K+, Ca2+, or Cl− (see Figure 5.4). These proteins are receptors because their functioning depends on ligand binding. An example is the acetylcholine receptor, a ligand-gated sodium channel located in the cell membranes of skeletal muscle cells. Acetylcholine is a neurotransmitter—a chemical signal released from nerve cells. Opening of the channel allows Na+, which is more concentrated outside the cell than inside, to diffuse into the cell. This initiates a series of events that result in muscle contraction (see Figure 34.9).
Protein Kinase Receptors
Like ligand-gated channel receptors, protein kinase receptors change shape upon ligand binding. But in this case, the new conformation exposes or activates a catalytic domain on the cytoplasmic side of the transmembrane protein that has protein kinase activity—it modifies specific target proteins in the cell by adding phosphate groups to them. In general, protein kinases catalyze the following reaction:
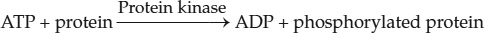
This reaction results in the covalent modification (phosphorylation) of the target protein, thereby changing its activity (see Figure 3.20B). Protein kinases are extraordinarily important in biological signaling: about 1 human gene in 50 is a protein kinase gene, and there is an even higher proportion of such genes in some plants.
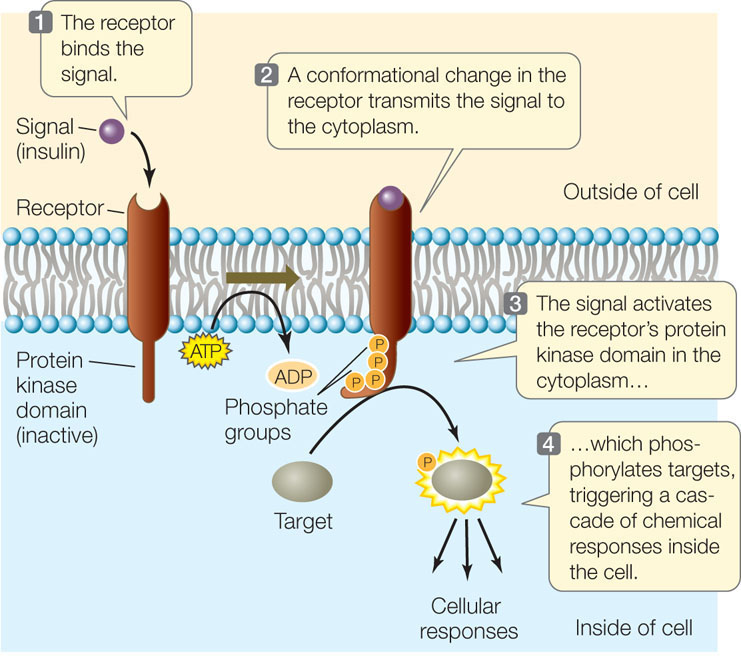
An example of a protein kinase receptor is the receptor for the hormone insulin (FIGURE 5.13). The activation of this receptor results in the phosphorylation of target proteins, which then bring about the cell’s response, which includes the insertion of glucose transport proteins into the cell membrane.
It should be noted that not all protein kinases are receptors—many function in later steps of signal transduction pathways, as we will discuss in Concept 5.6.
98
G Protein-Linked Receptors
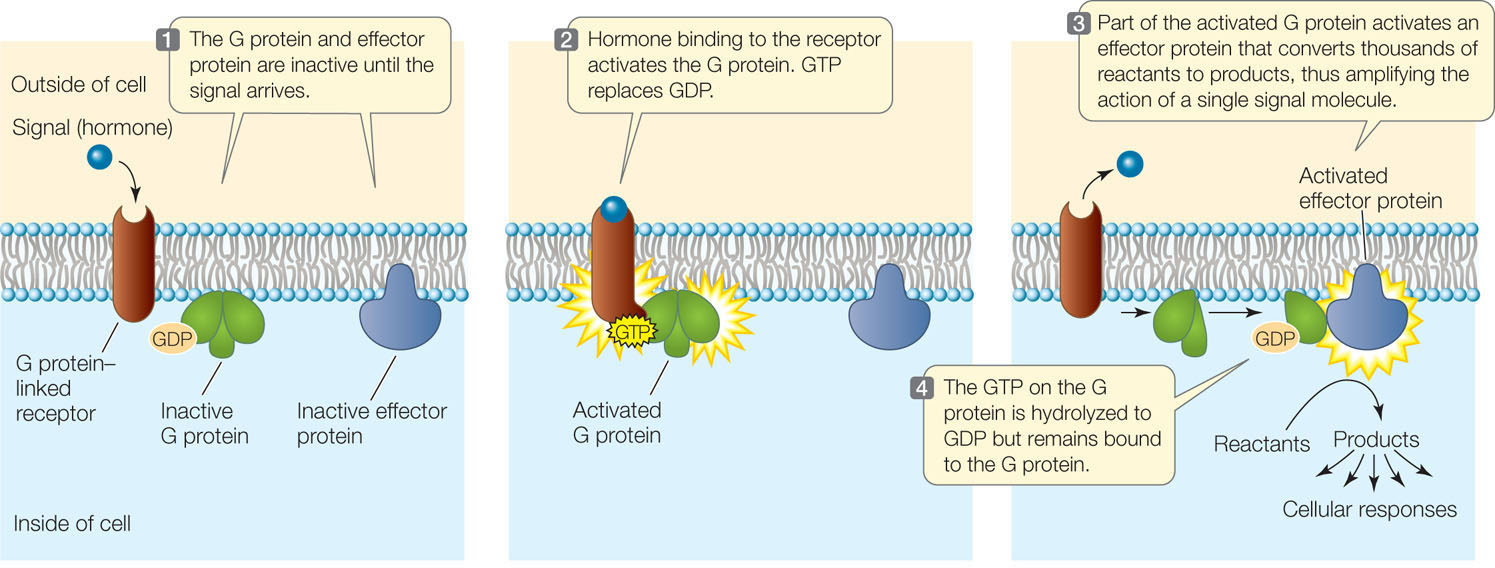
A third category of eukaryotic cell membrane receptors is the family of G protein–linked receptors. In this case, ligand binding on the extracellular domain of the receptor exposes a site on the cytoplasmic side that can bind to a mobile membrane protein called a G protein. The G protein is partially inserted in the lipid bilayer and partially exposed on the cytoplasmic surface of the membrane.
Many G proteins have three polypeptide subunits and can bind three different molecules:
- The G protein–linked receptor
- GDP and GTP (guanosine diphosphate and guanosine triphosphate; these are nucleotides, like ADP and ATP)
- An effector protein (a protein that causes an effect in the cell)
The activated G protein–linked receptor functions as a guanine nucleotide exchange factor. It exchanges a GDP nucleotide bound to the G protein for a GTP, inducing a shape change in the G protein. The G protein then activates the effector protein, leading to downstream signal amplification (FIGURE 5.14). G protein-linked receptors are especially important in the sensory systems of animals (see Concept 34.4).

Go to ANIMATED TUTORIAL 5.5 G Protein-Linked Signal Transduction and Cancer
PoL2e.com/at5.5
CHECKpoint CONCEPT 5.5
- Name three major steps in cell signaling that were discussed in this concept.
- What are the differences and similarities between ion channel receptors and G protein-linked receptors?
- If an intact cell is treated to remove cell surface proteins, will the cell be able to receive any environmental signals? Explain.
When a signal activates a receptor, a signal transduction pathway ensues. This often involves multiple steps, and it leads to one or more specific cellular responses. We will discuss these processes next.
99