Concept 19.2: Prokaryote Diversity Reflects the Ancient Origins of Life
The prokaryotes were alone on Earth for a very long time, adapting to new environments and to changes in existing environments. They have survived to this day, in massive numbers and incredible diversity, and they are found everywhere. In numbers of individuals, prokaryotes are far more abundant than eukaryotes are. Individual bacteria and archaea in the oceans number more than 3 × 1028—about a million times more than the number of stars in the universe. Closer to home, the individual bacteria living in your intestinal tract outnumber all the humans who have ever lived.
Given our still fragmentary knowledge of prokaryote diversity, it is not surprising that there are several different hypotheses about the relationships of the major groups of prokaryotes. In this book we use a widely accepted classification system that has considerable support from nucleotide sequence data. We will discuss eight major bacterial groups that have the broadest phylogenetic support and have received the most study: the low-GC Gram-positives, high-GC Gram-positives, hyperthermophilic bacteria, hadobacteria, cyanobacteria, spirochetes, chlamydias, and proteobacteria (see Figure 19.1). We will then describe the archaea, whose diversity is even less well studied than that of the bacteria.
384
The low-GC Gram-positives include the smallest cellular organisms
The low-GC Gram-positives, also known as Firmicutes, derive the first part of their name from the relatively low ratio of G-C to A-T nucleotide base pairs in their DNA. The second part of their name is less accurate: some of the low-GC Gram-positives are in fact Gram-negative, and some have no cell wall at all. Despite these differences, phylogenetic analyses of DNA sequences support the monophyly of this bacterial clade.
One group of low-GC Gram-positives can produce heat-resistant resting structures called endospores (FIGURE 19.5). When a key nutrient such as nitrogen or carbon becomes scarce, the bacterium replicates its DNA and encapsulates one copy, along with some of its cytoplasm, in a tough cell wall heavily thickened with peptidoglycan and surrounded by a spore coat. The parent cell then breaks down, releasing the endospore. Endospore production is not a reproductive process, as the endospore merely replaces the parent cell. The endospore, however, can survive harsh environmental conditions that would kill the parent cell, such as high or low temperatures or drought, because it is dormant—its normal metabolic activity is suspended. Later, if it encounters favorable conditions, the endospore becomes metabolically active and divides, forming new cells that are like the parent cells. Members of this endospore-forming group of low-GC Gram-positives include the many species of Clostridium and Bacillus. Some of their endospores can be reactivated after more than 1,000 years of dormancy. There are even credible claims of reactivation of Bacillus endospores that are millions of years old.
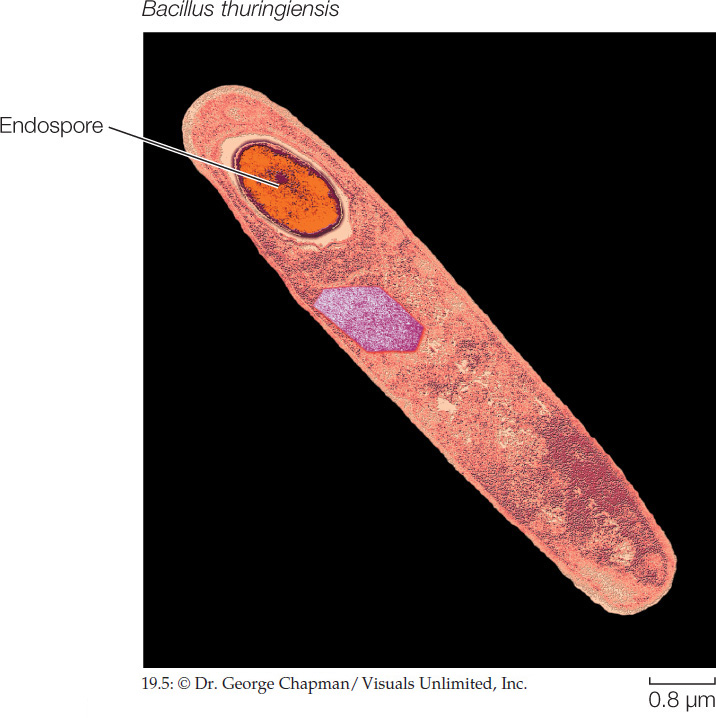
Endospores of Bacillus anthracis are the cause of anthrax. Anthrax is primarily a disease of grazing animals, but it can be fatal in humans if the infection starts in the respiratory system. When the endospores sense macrophages in mammalian blood, they reactivate and release toxins into the bloodstream. Bacillus anthracis has been used as a bioterrorism agent because large quantities of its endospores can be transported in a small space and then spread among human populations, where they may be inhaled or ingested.
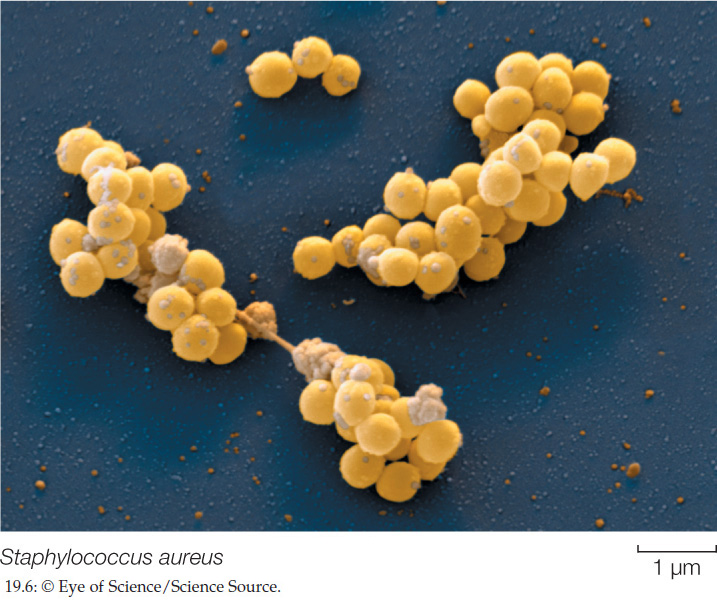
Low-GC Gram-positives of the genus Staphylococcus—the staphylococci (FIGURE 19.6)—are abundant on the human body surface; they are responsible for boils and many other skin problems. Staphylococcus aureus is the best-known human pathogen in this genus; it is present in 20–40 percent of normal adults (and in 50–70 percent of hospitalized adults). In addition to skin diseases, S. aureus can cause respiratory, intestinal, and wound infections.
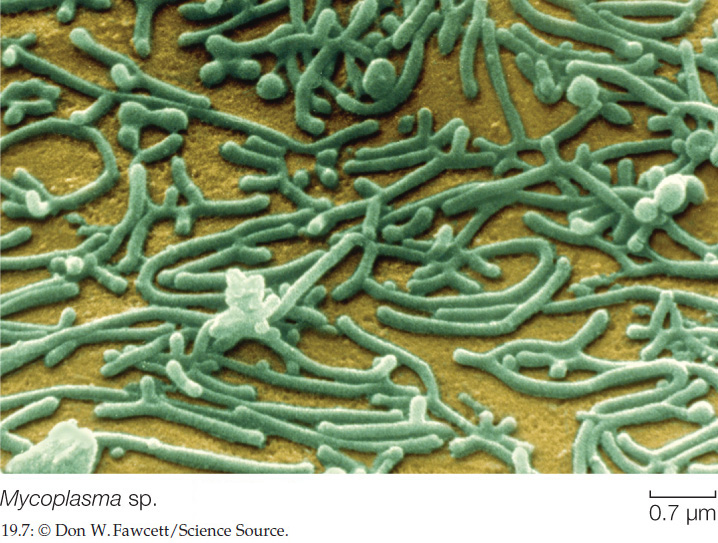
Another interesting group of low-GC Gram-positives, the mycoplasmas, lack cell walls, although some have a stiffening material outside the cell membrane. The mycoplasmas are among the smallest cellular organisms known (FIGURE 19.7). The smallest mycoplasmas capable of multiplying by binary fission have a diameter of about 0.2 μm. They are small in another crucial sense as well: they have less than half as much DNA as most other prokaryotes. It has been speculated that the DNA in a mycoplasma, which codes for fewer than 500 proteins, may be close to the minimum amount required to encode the essential properties of a living cell.
Some high-GC Gram-positives are valuable sources of antibiotics
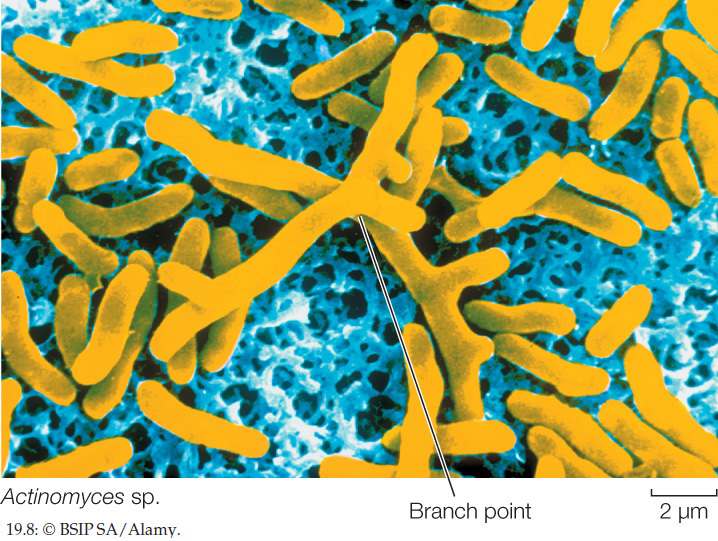
High-GC Gram-positives, also known as actinobacteria, have a higher ratio of G-C to A-T nucleotide base pairs than do the low-GC Gram-positives. These bacteria develop an elaborately branched system of filaments (FIGURE 19.8) that resembles the filamentous growth habit of fungi, albeit at a reduced scale. Some high-GC Gram-positives reproduce by forming chains of spores at the tips of the filaments. In species that do not form spores, the branched, filamentous growth ceases and the structure breaks up into typical cocci or bacilli, which then reproduce by binary fission.
385
The high-GC Gram-positives include several medically important bacteria. Mycobacterium tuberculosis causes tuberculosis, which kills 3 million people each year. Genetic data suggest that this bacterium arose 3 million years ago in East Africa, making it the oldest known human bacterial pathogen. The genus Streptomyces produces streptomycin as well as hundreds of other antibiotics. We derive most of our antibiotics from members of the high-GC Gram-positives.
Hyperthermophilic bacteria live at very high temperatures
Several lineages of bacteria and archaea are extremophiles: they thrive under extreme conditions that would kill most other organisms. The hyperthermophilic bacteria, for example, are thermophiles (Greek, “heat-lovers”). Genera such as Aquifex live near volcanic vents and in hot springs, sometimes at temperatures near the boiling point of water. Some species of Aquifex need only hydrogen, oxygen, carbon dioxide, and mineral salts to live and grow. Species of the genus Thermotoga live deep underground in oil reservoirs, as well as in other high-temperature environments.
Biologists have hypothesized that high temperatures characterized the ancestral conditions for life on Earth, given that most environments on early Earth were much hotter than those of today. Reconstructions of ancestral bacterial genes have supported this hypothesis by showing that the ancestral sequences functioned best at elevated temperatures. The monophyly of the hyperthermophilic bacteria, however, is not well established.
Hadobacteria live in extreme environments
The hadobacteria, including such genera as Deinococcus and Thermus, are another group of thermophilic extremophiles. The group’s name is derived from Hades, the ancient Greek name for the underworld. Deinococcus are resistant to radiation and can consume nuclear waste and other toxic materials. They can also survive extremes of cold as well as hot temperatures. An other member of this group, Thermus aquaticus, was the source of the thermally stable DNA polymerase that was critical for the development of the polymerase chain reaction. Thermus aquaticus was originally isolated from a hot spring, but it can of be found wherever hot water occurs (including many residential hot water heaters).
Cyanobacteria were the first photosynthesizers
Cyanobacteria, sometimes called blue-green bacteria because of their pigmentation, are photosynthetic bacteria that require only water, nitrogen gas, oxygen, a few mineral elements, light, and carbon dioxide to survive. They use chlorophyll a for photosynthesis and release oxygen gas (O2); many species also fix nitrogen. Photosynthesis by these bacteria was the basis of the “green revolution” that transformed Earth’s atmosphere (see Concept 18.3).
Cyanobacteria carry out the same type of photosynthesis that is characteristic of eukaryotic photosynthesizers. They contain elaborate and highly organized internal membrane systems called photosynthetic lamellae. As mentioned in Concept 19.1, the chloroplasts of photosynthetic eukaryotes are derived from an endosymbiotic cyanobacterium.
Cyanobacteria may live free as single cells or associate in multicellular colonies. Depending on the species and on growth conditions, these colonies may range from flat sheets one cell thick to filaments to spherical balls of cells. Some filamentous colonies of cyanobacteria differentiate into three specialized cell types: vegetative cells, spores, and heterocysts. Vegetative cells photosynthesize, spores are resting stages that can survive harsh environmental conditions and eventually develop into new filaments, and heterocysts are cells specialized for nitrogen fixation (FIGURE 19.9). All of the known cyanobacteria with heterocysts fix nitrogen. Heterocysts also have a role in reproduction: when filaments break apart to reproduce, the heterocyst may serve as a breaking point.


Go to MEDIA CLIP 19.1 Cyanobacteria
PoL2e.com/mc19.1
386
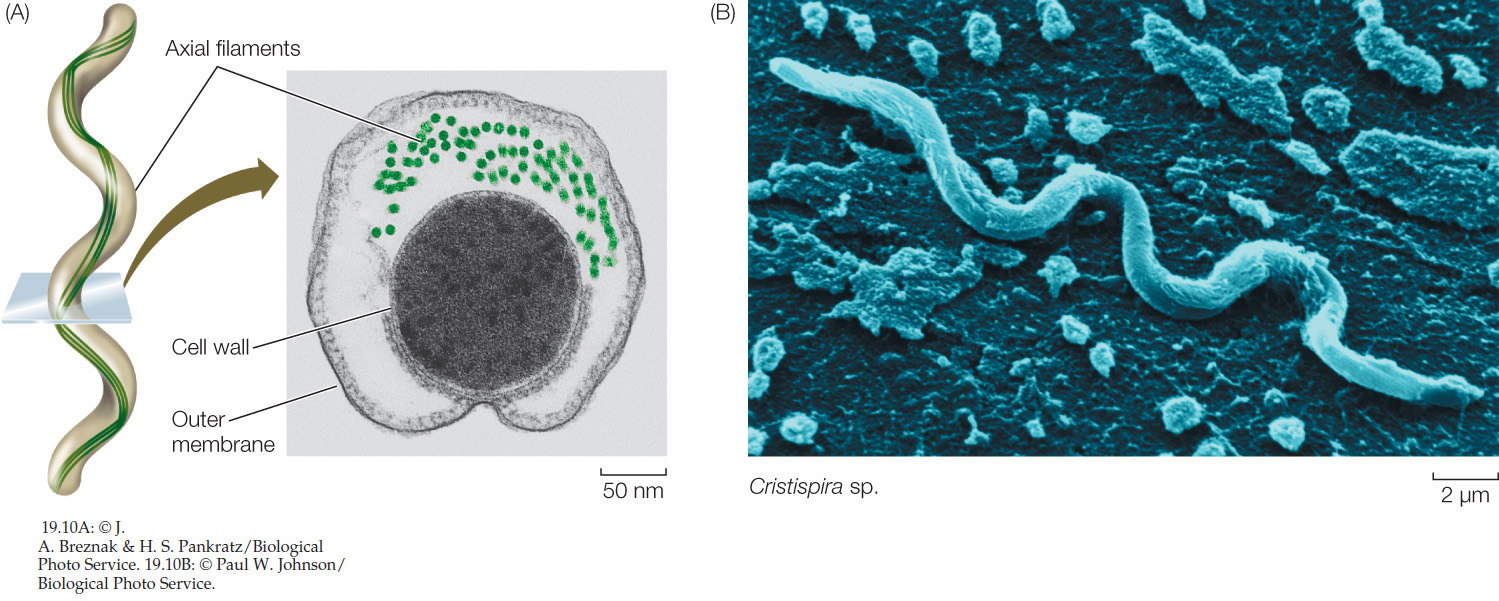
Spirochetes move by means of axial filaments
Spirochetes are Gram-negative, motile bacteria characterized by unique structures called axial filaments (FIGURE 19.10A), which are modified flagella running through the periplasmic space. The cell body is a long cylinder coiled into a spiral (FIGURE 19.10B). The axial filaments begin at either end of the cell and overlap in the middle. Motor proteins connect the axial filaments to the cell wall, enabling the corkscrew-like movement of the bacterium. Many spirochetes are parasites of humans; a few are pathogens, including those that cause syphilis and Lyme disease. Others live free in mud or water.
Chlamydias are extremely small parasites
Chlamydias are among the smallest bacteria (0.2–1.5 μm in diameter). They can live only as parasites in the cells of other organisms. It was once believed that their obligate parasitism resulted from an inability to produce ATP—that chlamydias were “energy parasites.” However, genome sequencing indicates that chlamydias have the genetic capacity to produce at least some ATP. They can augment this capacity by using an enzyme called a translocase, which allows them to take up ATP from the cytoplasm of their host in exchange for ADP from their own cells.
387
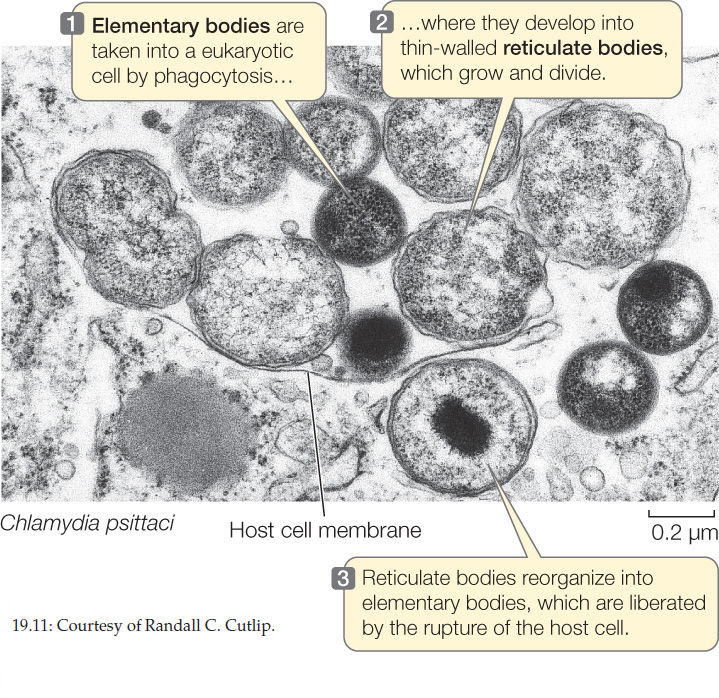
These tiny, Gram-negative cocci are unique among prokaryotes because of a complex life cycle that involves two different forms of cells, elementary bodies and reticulate bodies (FIGURE 19.11). Various strains of chlamydias cause eye infections (especially trachoma), sexually transmitted diseases, and some forms of pneumonia in humans.
The proteobacteria are a large and diverse group
By far the largest group of bacteria, in terms of number of described species, is the proteobacteria. The proteobacteria include many species of Gram-negative photoautotrophs that use light-driven reactions to metabolize sulfur, as well as dramatically diverse bacteria that bear no phenotypic resemblance to the photoautotrophic species. Genetic and morphological evidence indicates that the mitochondria of eukaryotes were derived from a proteobacterium by endosymbiosis.
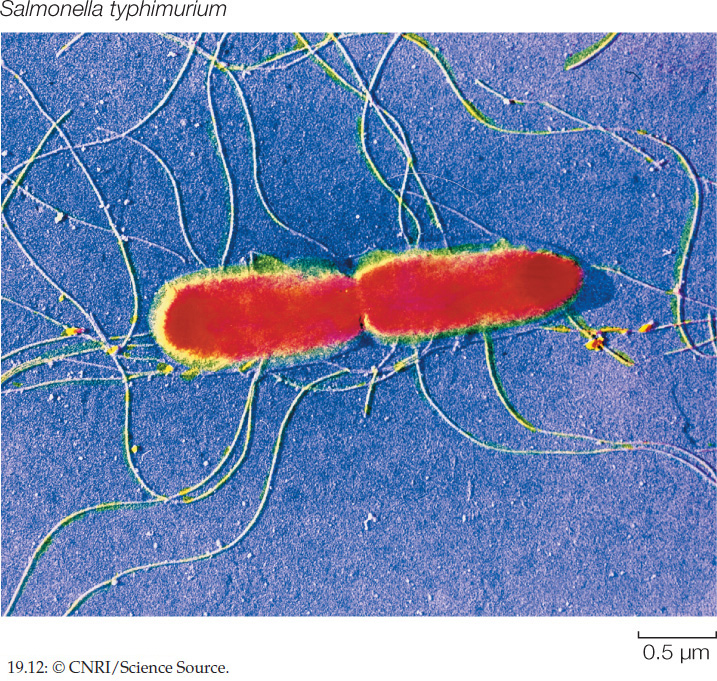
Among the proteobacteria are some nitrogen-fixing genera, such as Rhizobium, and other bacteria that contribute to the global nitrogen and sulfur cycles. Escherichia coli, one of the most studied organisms on Earth, is a proteobacterium. So, too, are many of the most famous human pathogens, such as Yersinia pestis (which causes bubonic plague), Vibrio cholerae (cholera), and Salmonella typhimurium (gastrointestinal disease; FIGURE 19.12).

Go to MEDIA CLIP 19.2 A Swarm of Salmonella
PoL2e.com/mc19.2
The bioluminescent Vibrio we discussed at the opening of this chapter are also members of this group. There are many potential applications of the genes that encode bioluminescent proteins in bacteria. Already, these genes are being inserted into the genomes of other species in which the resulting bioluminescence is used as a marker of gene expression. Futuristic proposals for making use of bioluminescence in bioengineered organisms include crop plants that glow when they become water-stressed and need to be irrigated, and glowing trees that could light highways at night in place of electric lights.
Although fungi cause most plant diseases, and viruses cause others, about 200 known plant diseases are of bacterial origin. Crown gall, with its characteristic tumors (FIGURE 19.13), is one of the most striking. The causal agent of crown gall is Agrobacterium tumefaciens, a proteobacterium that harbors a plasmid used in recombinant DNA studies as a vehicle for inserting genes into new plant hosts.

Gene sequencing enabled biologists to differentiate the domain Archaea
The separation of Archaea from Bacteria and Eukarya was originally based on phylogenetic relationships determined from sequences of rRNA genes. It was supported when biologists sequenced the first archaeal genome. That genome consisted of 1,738 genes, more than half of which were unlike any genes ever found in the other two domains.
388
Archaea are well known for living in extreme habitats such as those with high salinity (salt content), low oxygen concentrations, high temperatures, or high or low pH (FIGURE 19.14). Many archaea are not extremophiles, however, but live in moderate habitats—they are common in soil, for example. Perhaps the largest numbers of archaea live in the ocean depths.
Investigation
HYPOTHESIS
Some prokaryotes can survive at temperatures above the 120°C threshold of sterilization.
METHOD
- Seal samples of unidentified, iron-reducing, thermal vent prokaryotes in tubes with a medium containing Fe3+ as an electron acceptor. Control tubes contain Fe3+ but no organisms.
- Hold both tubes in a sterilizer at 121°C for 10 hours. If the iron-reducing organisms are metabolically active, they will reduce the Fe3+ to Fe2+ (as magnetite, which can be detected with a magnet).
RESULTS

CONCLUSION
This thermal vent organism (Strain 121) can survive at temperatures above the previously defined sterilization limit.
ANALYZE THE DATA
After Strain 121 was isolated, its growth was examined at various temperatures. The table shows generation time (the time between cell divisions) at nine temperatures.
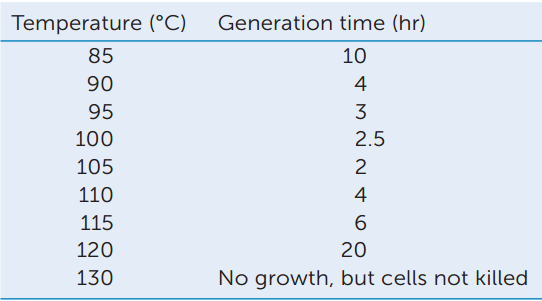
- Make a graph showing generation time as a function of temperature.
- Which temperature appears to be closest to the optimum for growth of Strain 121?
- Note that no growth occurred at 130°C, but that the cells were not killed. How would you demonstrate that these cells were still alive?
Go to LaunchPad for discussion and relevant links for all INVESTIGATION figures.
aK. Kashefi and D. R. Lovley. 2003. Science 301: 934.
One current classification scheme divides Archaea into two principal groups, Crenarchaeota and Euryarchaeota. Less is known about three more recently discovered groups, Korarchaeota, Nanoarchaeota, and Thaumarchaeota. In fact, we know relatively little about the phylogeny of archaea, in part because the study of these prokaryotes is still in its early stages.
Two characteristics shared by all archaea are the absence of peptidoglycan in their cell walls and the presence of lipids of distinctive composition in their cell membranes (see Table 19.1). The unusual lipids in the membranes of archaea are found in all archaea and in no bacteria or eukaryotes. Most lipids in bacterial and eukaryotic membranes contain unbranched long-chain fatty acids connected to glycerol molecules by ester linkages:
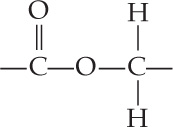
In contrast, some lipids in archaeal membranes contain long-chain hydrocarbons connected to glycerol molecules by ether linkages:
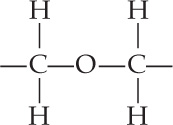
389
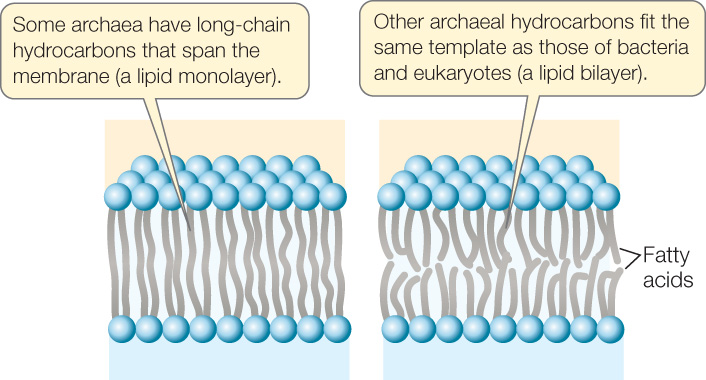
These ether linkages are a synapomorphy (a shared, derived trait viewed as evidence of common ancestry) of archaea. In addition, the long-chain hydrocarbons of archaea are branched. One class of archaeal lipids, with hydrocarbon chains 40 carbon atoms in length, contains glycerol at both ends of the hydrocarbons (FIGURE 19.15). These lipids form a lipid monolayer structure that is unique to archaea. They still fit into a biological membrane because they are twice as long as the typical lipids in the bilayers of other membranes. Lipid monolayers and bilayers are both found among the archaea. The effects, if any, of these structural features on membrane performance are unknown. In spite of this striking difference in their lipids, the membranes of all three domains have similar overall structures, dimensions, and functions.
Most crenarchaeotes live in hot or acidic places
Most known crenarchaeotes are either thermophilic, acidophilic (acid loving), or both. Members of the genus Sulfolobus live in hot sulfur springs at temperatures of 70°C–75°C. They become metabolically inactive at 55°C (131°F). Hot sulfur springs are also extremely acidic. Sulfolobus grows best in the range of pH 2–pH 3, but some members of this genus readily tolerate pH values as low as 0.9. Most acidophilic thermophiles maintain an internal pH of 5.5–7 (close to neutral) in spite of their acidic environment. These and other thermophiles thrive where few other organisms can even survive (FIGURE 19.16).

Euryarchaeotes are found in surprising places
Some species of Euryarchaeota are methanogens: they produce methane (CH4) by reducing carbon dioxide, and this is the key step in their energy metabolism. All of the methanogens are obligate anaerobes (see Concept 19.3). Comparison of their rRNA gene sequences has revealed a close evolutionary relationship among these methanogenic species, which were previously assigned to several different groups of bacteria.
Methanogenic euryarchaeotes release approximately 2 billion tons of methane gas into Earth’s atmosphere each year, accounting for 80–90 percent of the methane that enters the atmosphere, including that produced in some mammalian digestive systems. Approximately a third of this methane comes from methanogens living in the guts of ruminants such as cattle, sheep, and deer, and another large fraction comes from methanogens living in the guts of termites and cockroaches. Methane is increasing in Earth’s atmosphere by about 1 percent per year and contributes to the greenhouse effect (see Concept 45.4). Part of that increase is due to increases in cattle and rice farming and the methanogens associated with both.
Another group of euryarchaeotes, the extreme halophiles (salt lovers), lives exclusively in very salty environments. Because they contain pink carotenoid pigments, these archaea are sometimes easy to see (FIGURE 19.17). Extreme halophiles grow in the Dead Sea and in brines of all types. The reddish pink spots that can occur on pickled fish are colonies of halophilic archaea. Few other organisms can live in the saltiest of the homes that the extreme halophiles occupy. In these environments, most organisms would “dry” to death, losing too much water to the hypertonic environment. Extreme halophiles have been found in lakes with pH values as high as 11.5—the most alkaline environment inhabited by living organisms, and almost as alkaline as household ammonia.

390
Some of the extreme halophiles have a unique system for trapping light energy and using it to form ATP—without using any form of chlorophyll—when oxygen is in short supply. They use the pigment retinal (also found in the vertebrate eye) combined with a protein to form a light-absorbing molecule called microbial rhodopsin.
Another member of the Euryarchaeota, Thermoplasma, has no cell wall. It is thermophilic and acidophilic, its metabolism is aerobic, and it lives in coal deposits. Its genome of 1,100,000 base pairs is among the smallest (along with the mycoplasmas) of any free-living organism, although some parasitic organisms have even smaller genomes.
Several lineages of Archaea are poorly known
Most known archaea are crenarchaeotes or euryarchaeotes, but studies of extreme environments have identified several small lineages that are not closely related to either of these major groups. For example, the korarchaeotes and thaumarchaeotes are known only from DNA isolated directly from hot environments. Neither group has been successfully grown in pure culture. The thaumarchaeotes oxidize ammonia and may play an important role in the nitrogen cycle.
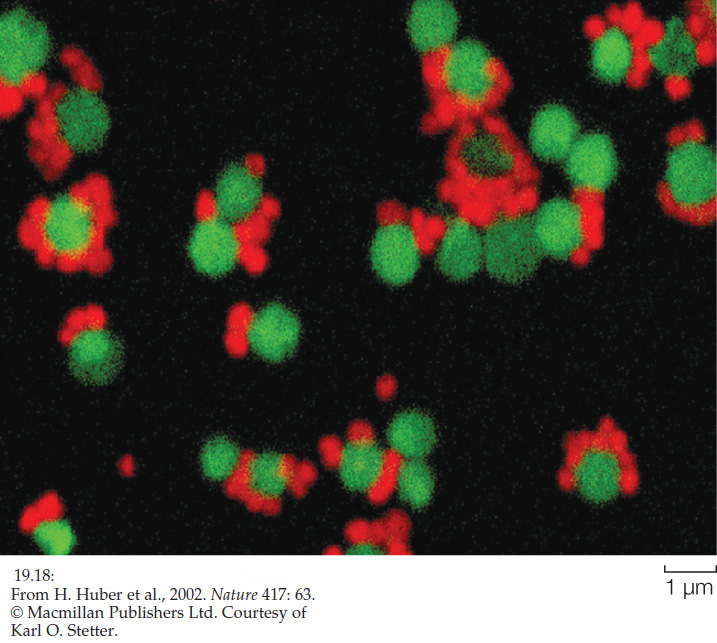
Another distinctive archaeal lineage has been discovered at a deep-sea hydrothermal vent off the coast of Iceland. It is the first representative of the group christened Nanoarchaeota because of their minute size (Greek nanos, “dwarf”). This organism is a parasite that lives on cells of Ignicoccus, a crenarchaeote. Because of their association, the two species can be grown together in culture (FIGURE 19.18).
CHECKpoint CONCEPT 19.2
- How does the diversity of environments occupied by prokaryotes compare with the diversity of environments occupied by multicellular organisms you may be familiar with?
- Explain why Gram staining is of limited use in understanding the evolutionary relationships of bacteria.
- What makes the membranes of archaea unique?
- Given that all species of life have evolved for the same amount of time since their common origin, how would you respond to someone who characterizes prokaryotes as “primitive”?
Prokaryotes are found almost everywhere on Earth and live in a wide variety of ecosystems. Next we will examine the contributions of prokaryotes to the functioning of those ecosystems.