Concept 22.2: Fungi Can Be Saprobic, Parasitic, Predatory, or Mutualistic
Without the fungi, our planet would be very different. Picture Earth with only a few stunted plants and watery environments choked with the remains of dead organisms. Fungi do much of Earth’s garbage disposal. Fungi not only help clean up the landscape and form soil, but also play a key role in recycling mineral nutrients. Furthermore, the colonization of the terrestrial environment was made possible in large part by associations that fungi formed with land plants and other organisms.
453
Saprobic fungi are critical to the planetary carbon cycle
Saprobic fungi, along with bacteria, are the major decomposers on Earth, contributing to the decay of nonliving organic matter and thus to recycling of the elements used by living things. In forests, for example, the mycelia of fungi absorb nutrients from fallen trees, thus decomposing their wood. Fungi are the principal decomposers of cellulose and lignin, the main components of plant cell walls (most bacteria cannot break down these materials). Other fungi produce enzymes that decompose keratin and thus break down animal structures such as hair and nails.
Were it not for the fungal decomposers, Earth’s carbon cycle would fail. Great quantities of carbon atoms would remain trapped forever on forest floors and elsewhere. Instead, those carbon atoms are returned to the atmosphere in the form of CO2 by fungal respiration, where they are again available for photosynthesis by plants.
LINK
Earth’s carbon cycle is described in Concept 45.3
There was a time in Earth’s history when populations of saprobic fungi declined dramatically. Vast tropical swamps existed during the Carboniferous period, as we saw in Chapter 21. When plants in these swamps died, they began to form peat. Peat formation led to acidification of the swamps. That acidity, in turn, drastically reduced the fungal population. The result? With the decomposers largely absent, large quantities of peat remained on the swamp floor and over time were converted into coal.
In contrast to their decline during the Carboniferous, fungi did very well at the end of the Permian, a quarter of a billion years ago, when the aggregation of continents produced volcanic eruptions that triggered a global mass extinction (see Chapter 18). The fossil record shows that even as 96 percent of all multicellular species became extinct, fungi flourished—demonstrating both their hardiness and their role in recycling the elements in dead plants and animals.
Simple sugars and the breakdown products of complex polysaccharides are the favored source of carbon for saprobic fungi. Most fungi obtain nitrogen from proteins or the products of protein breakdown. Many fungi can use nitrate (NO3−) or ammonium (NH4+) ions as their sole source of nitrogen. No known fungus can get its nitrogen directly from inorganic nitrogen gas, however, as can some bacteria and plant–bacteria associations (that is, fungi cannot fix nitrogen; see Concept 19.3).
What happens when a fungus faces a dwindling food supply? A common strategy is to reproduce rapidly and abundantly. When conditions are good, fungi produce great quantities of spores, but the rate of spore production is commonly even higher when nutrient supplies go down. The spores may then remain dormant until conditions improve, or they may be dispersed to areas where nutrient supplies are higher.
Not only are fungal spores abundant in number, but they are extremely tiny and easily spread by wind or water (FIGURE 22.4). These attributes virtually ensure that they will be scattered over great distances, and that at least some of them will find conditions suitable for growth. The air we breathe contains as many as 10,000 fungal spores per cubic meter. No wonder we find fungi just about everywhere.


Go to MEDIA CLIP 22.1 Fungal Decomposers
PoL2e.com/mc22.1
Some fungi engage in parasitic or predatory interactions
Whereas saprobic fungi obtain their energy, carbon, and nitrogen directly from dead organic matter, other species of fungi obtain their nutrition from parasitic—and even predatory—interactions.
Parasitic Fungi
Mycologists (biologists who study fungi) distinguish between two classes of parasitic fungi based on their degree of dependence on their host. Facultative parasites can grow on living organisms but can also grow independently (including on artificial media). Obligate parasites can grow only on a living host. The fact that their growth depends on a living host shows that obligate parasites have specialized nutritional requirements.
Plants and insects are the most common hosts of parasitic fungi. The filamentous structure of fungal hyphae is especially well suited to a life of absorbing nutrients from living plants. The slender hyphae of a parasitic fungus can invade a plant through stomata, through wounds, or in some cases, by direct penetration of epidermal cell walls (FIGURE 22.5A). Once inside the plant, the hyphae branch out to expand the mycelium. Some hyphae produce haustoria (singular haustorium), branching projections that push through cell walls into living plant cells, absorbing the nutrients within those cells. The haustoria do not break through the cell membranes inside the cell walls, but instead push pockets into the membranes, so that the cell membrane fits them like a glove (FIGURE 22.5B). Fruiting structures may form, either within the plant body or on its surface.
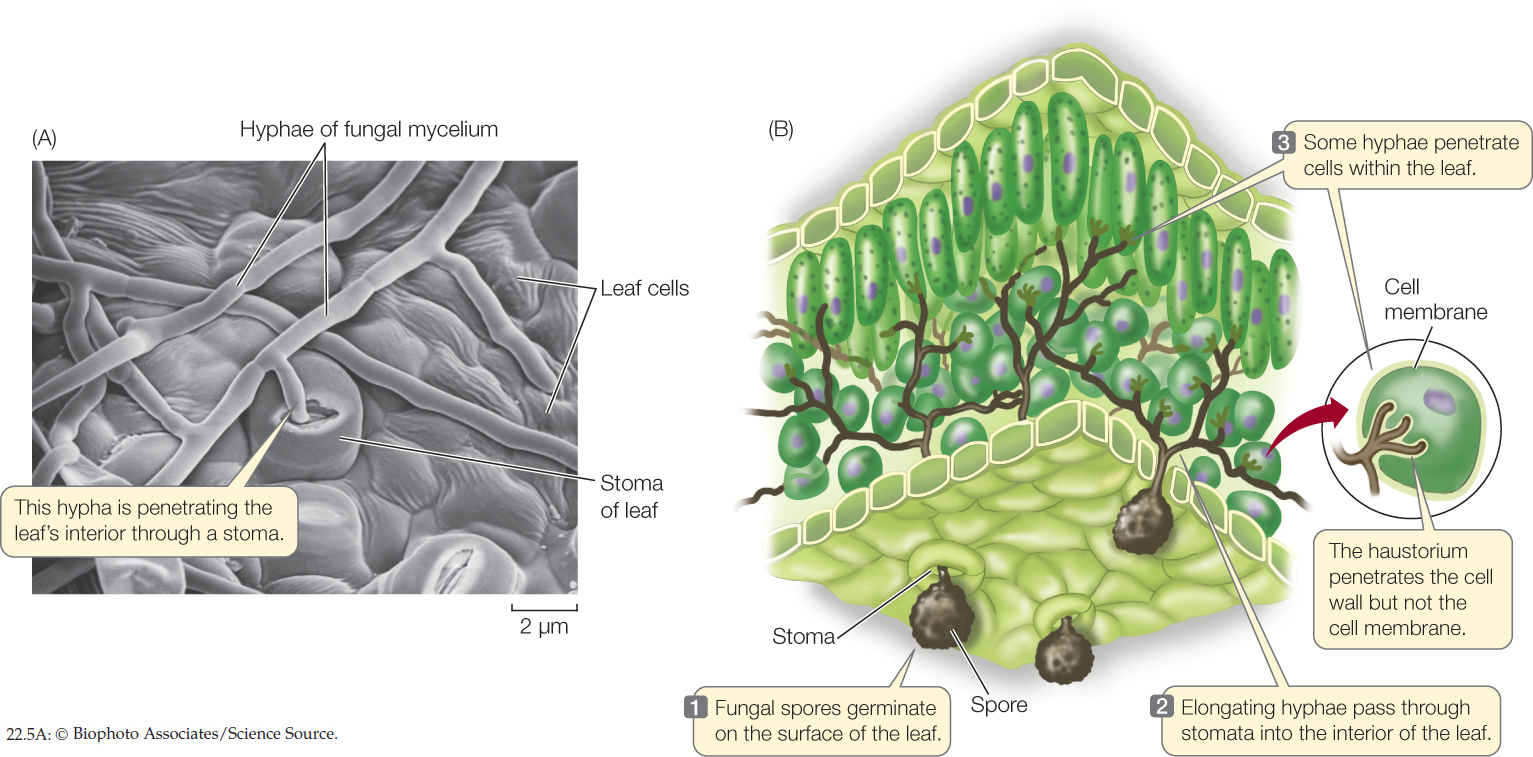
454
Some parasitic fungi live in a close physical (symbiotic) relationship with their host that is usually not lethal to the plant. Others are pathogenic, sickening or even killing the host from which they derive nutrition.

Go to MEDIA CLIP 22.2 Mind-Control Killer Fungi
PoL2e.com/mc22.2
Pathogenic Fungi
Although most human diseases are caused by bacteria or viruses, fungal pathogens are a major cause of death among people with compromised immune systems. Many people with AIDS die of fungal diseases, such as the pneumonia caused by Pneumocystis jirovecii. Even Candida albicans and certain other yeasts that are normally a part of a healthy microbiome can cause severe diseases, such as esophagitis (which impairs swallowing), in individuals with AIDS and in individuals taking immunosuppressive drugs. Various fungi cause other, less threatening human diseases, such as ringworm and athlete’s foot. Our limited understanding of the basic biology of these fungi still hampers our ability to treat the diseases they cause. As a result, fungal diseases are a growing international health problem.
The worldwide decline of amphibian species has been linked to the spread of a chytrid fungus, Batrachochytrium dendrobatidis (or Bd for simplicity). In some areas of the world where this fungus has been present for millennia, amphibian populations appear to be tolerant of Bd and it does not cause widespread die-offs. But in other areas, such as western North America and Australia, introductions of Bd are implicated in the loss or decline of many frog and salamander populations, or the extinction of entire species. Some strains of Bd are native to southern Africa, where they do not appear to cause widespread amphibian declines. Biologists have hypothesized that the spread of Bd around the world may have been initiated in the 1930s with exports of the African clawed frog (Xenopus laevis), which was once widely used in human pregnancy tests. Recent studies by Erica Rosenblum and her colleagues have shown that Africa may indeed have been one source of the introductions, but that strains of Bd appear to have been introduced multiple times from several geographic areas (FIGURE 22.6).
Investigation
HYPOTHESIS
Pathogenic forms of Bd that kill many amphibians were introduced into western North America from Africa, through importation of African clawed frogs beginning in the 1930s.
METHOD
- Collect samples of Bd from throughout the world.
- Sequence the genomes of the sampled chytrids.
- Construct a phylogenetic tree by comparing the DNA sequences of the different samples of Bd.
- Assess whether or not the resulting tree is consistent with the introduction of Bd from Africa to western North America.
RESULTS
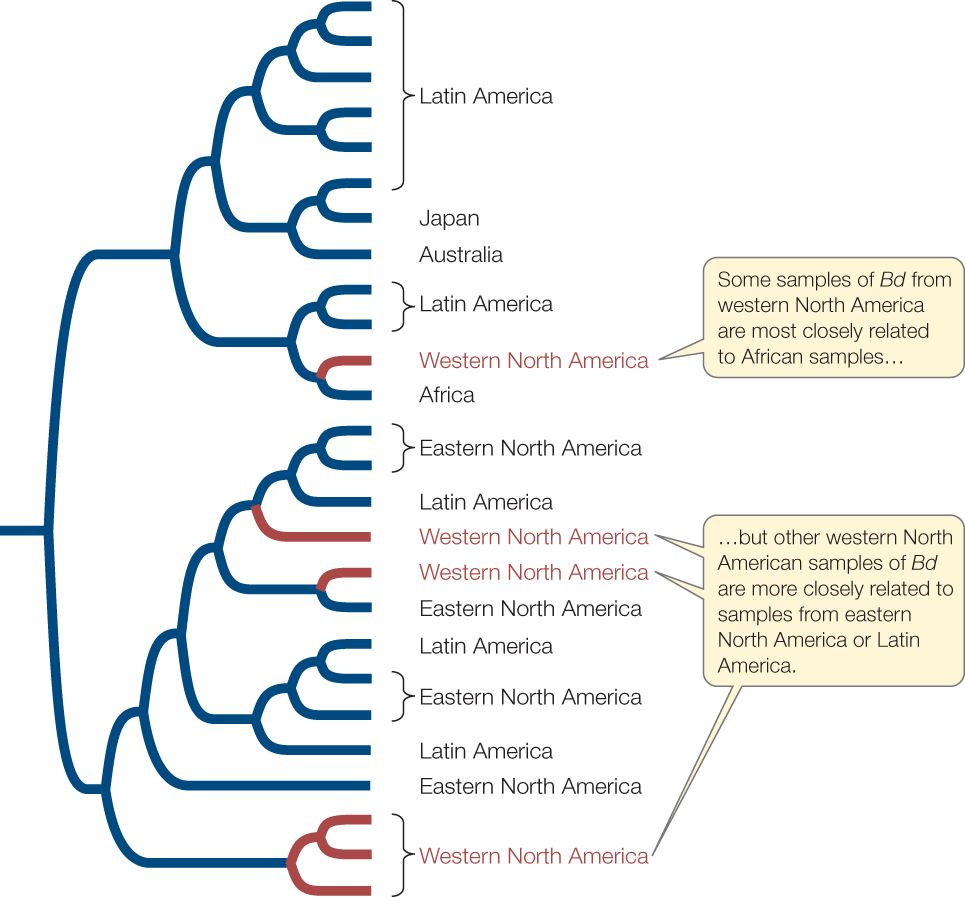
CONCLUSION
Pathogenic strains of Bd appear to have been introduced to western North America from several different sources. At least one strain of Bd is consistent with an introduction from Africa, but other strains were more likely introduced from eastern North America or Latin America.
Go to LaunchPad for discussion and relevant links for all INVESTIGATION figures.
aE. B. Rosenblum et al. 2013. Proceedings of the National Academy of Sciences USA 110: 9385–9390.
Fungi are by far the most important plant pathogens, causing crop losses amounting to billions of dollars. Bacteria and viruses are less important than fungi as plant pathogens. Major fungal diseases of crop plants include black stem rust of wheat and other diseases of wheat, corn, and oats. The agent of black stem rust is Puccinia graminis, which has a complicated life cycle that involves two plant hosts (wheat and barberry). In an epidemic in 1935, P. graminis was responsible for the loss of about one-fourth of the wheat crop in Canada and the United States.
Although many pathogenic fungi cause problems when they attack agricultural crops, pathogenic fungi of the genus Fusarium can benefit agriculture by killing certain weed species, such as witchweed, a serious pest of cereal crops. Another strain of Fusarium has been proposed as a tool in the war against cocaine. The fungus could be applied to kill coca plants, the source of the drug. The use of Fusarium to kill coca plants is highly controversial, however, as the fungus might infect neighboring plant species in addition to the coca plants.
Predatory Fungi
Some fungi have adaptations that enable them to function as active predators, trapping nearby microscopic protists or animals. The most common predatory strategy seen in fungi is to secrete sticky substances from the hyphae so that passing organisms stick to them. The hyphae then quickly invade the trapped prey, growing and branching within it, spreading through its body, absorbing nutrients, and eventually killing it.
455
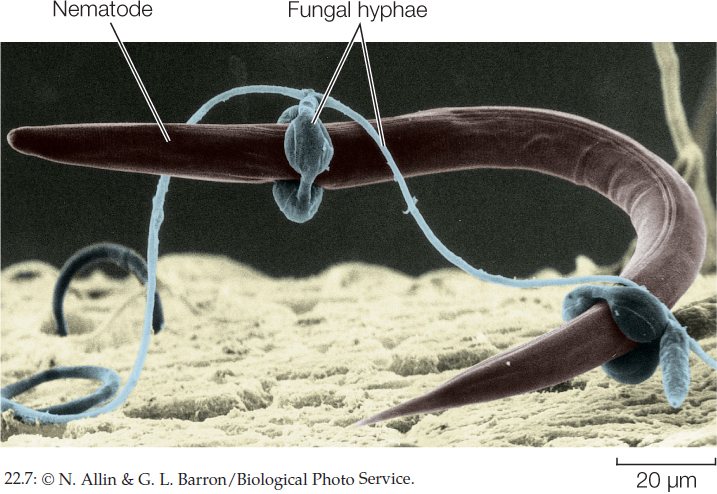
A more dramatic adaptation for predation is the constricting ring formed by some species of soil fungi (FIGURE 22.7). When nematodes (tiny roundworms) are present in the soil, these fungi form three-celled rings with a diameter that just fits a nematode. A nematode crawling through one of these rings stimulates the fungus, causing the cells of the ring to swell and trap the worm. Fungal hyphae quickly invade and digest the unlucky victim.
Mutualistic fungi engage in relationships beneficial to both partners
Certain kinds of relationships between fungi and other organisms have nutritional consequences for both partners. Two relationships of this type are highly specific and are symbiotic (the partners live in close, permanent contact with each other) as well as mutualistic (the relationship benefits both partners).
LINK
The obligate mutualistic relationship between leaf-cutter ants and the fungus they cultivate is necessary for the survival of each partner; see the opener of Chapter 43 and Concepts 43.3 and 43.4
Lichens
A lichen is not a single organism, but rather a symbiosis between two radically different species: a fungus and a photosynthetic microorganism. Together the organisms that constitute a lichen can survive some of the harshest environments on Earth (although they are sensitive to poor air quality; see Concept 22.4). The biota of Antarctica, for example, features more than 100 times as many species of lichens as of vascular plants. Relatively little experimental work has focused on lichens, perhaps because they grow so slowly—typically less than 1 centimeter in a year.
There are nearly 20,000 described “species” of lichens, each of which is assigned the name of its fungal component. These fungal components may constitute as many as 20 percent of all fungal species. Most of them are sac fungi (Ascomycota). Some of them are able to grow independently without a photosynthetic partner, but most have never been observed in nature other than in a lichen association. The photosynthetic component of a lichen is most often a unicellular green alga, but it can be a cyanobacterium, or may even include both.
456
Lichens are found in all sorts of exposed habitats: on tree bark, on open soil, and on bare rock. Reindeer moss (not a moss at all, but the lichen Cladonia subtenuis) covers vast areas in Arctic, sub-Arctic, and boreal regions, where it is an important part of the diets of reindeer and other large mammals.
The body forms of lichens fall into three principal categories: Crustose (crustlike) lichens look like colored powder dusted over their substrate (FIGURE 22.8A). Foliose (leafy) lichens are more loosely attached and have large leaflike forms, whereas fruticose (shrubby) lichens are highly branched and grow upward like little shrubs, or hang in long strands from tree branches or rocks (FIGURE 22.8B).

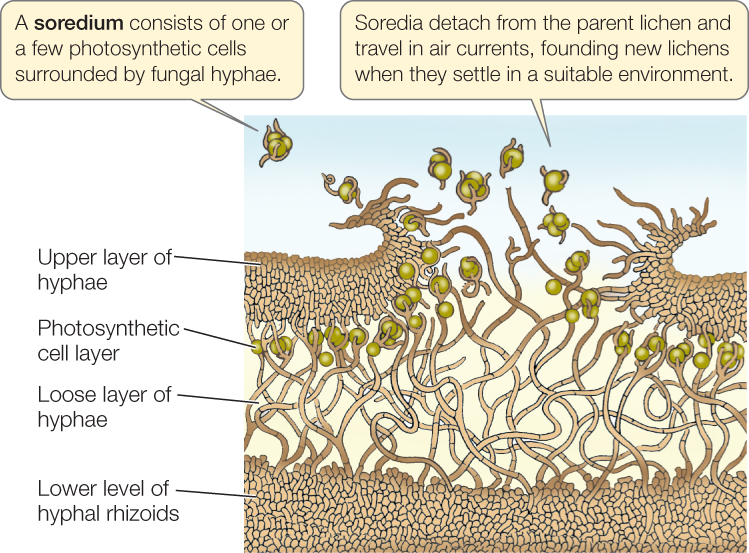
A cross section of a typical foliose lichen reveals a tight upper region of fungal hyphae, a layer of photosynthetic cyanobacteria or algae, a looser hyphal layer, and finally hyphal rhizoids that attach the entire structure to its substrate (FIGURE 22.9). The meshwork of fungal hyphae takes up some mineral nutrients needed by the photosynthetic cells and also holds water tenaciously, providing a suitably moist environment. The fungi obtain fixed carbon from the photosynthetic products of the algal or cyanobacterial cells.
Within the lichen, the fungal hyphae are tightly pressed against the algal or cyanobacterial cells and sometimes even invade them without breaching the cell membrane (similar to the haustoria in parasitic fungi; see Figure 22.5). The photosynthetic cells not only survive these intrusions but continue to grow. Algal cells in a lichen “leak” photosynthetic products at a greater rate than do similar cells growing on their own, and photosynthetic cells taken from lichens grow more rapidly on their own than when associated with a fungus. On the basis of these observations, we could consider lichen fungi to be parasitic on their photosynthetic partners. In many places where lichens grow, however, the photosynthetic cells could not grow at all on their own.
Lichens can reproduce simply by fragmentation of the vegetative body (the thallus) or by means of specialized structures called soredia (singular soredium). Soredia consist of one or a few photosynthetic cells bound by fungal hyphae. They become detached from the lichen, are dispersed by air currents, and upon arriving at a favorable location, develop into a new lichen thallus. Alternatively, the fungal partner may go through its sexual reproductive cycle, producing haploid spores. When these spores are discharged, however, they disperse alone, unaccompanied by the photosynthetic partner.
Lichens are often the first colonists on new areas of bare rock. They get most of the mineral nutrients they need from the air and rainwater, augmented by minerals absorbed from dust. A lichen begins to grow shortly after a rain, as it begins to dry. As it grows, the lichen acidifies its environment slightly, and this acidity contributes to the slow breakdown of rocks, an early step in soil formation. With further drying, the lichen’s photosynthesis ceases. The water content of the lichen may drop to less than 10 percent of its dry weight, at which point it becomes highly insensitive to extremes of temperature.
Mycorrhizae
Many vascular plants depend on a symbiotic association with fungi. This ancient association between plants and fungi was critical to the successful exploitation of the terrestrial environment by plants. Unassisted, the root hairs of many plants often do not take up enough water or minerals to sustain growth. However, the roots of such plants usually become infected with fungi, forming an association called a mycorrhiza (plural mycorrhizae). There are two types of mycorrhizae, distinguished by whether or not the fungal hyphae penetrate the plant cell walls.
457
In ectomycorrhizae, the fungus wraps around the root, and its mass is often as great as that of the root itself (FIGURE 22.10A). The fungal hyphae wrap around individual cells in the root but do not penetrate the cell walls. An extensive web of hyphae penetrates the soil in the area around the root, so that up to 25 percent of the soil volume near the root may be fungal hyphae. The hyphae attached to the root increase the surface area for the absorption of water and minerals, and the mass of hyphae in the soil acts like a sponge to hold water in the neighborhood of the root. Infected roots are short, swollen, and club-shaped, and they lack root hairs.
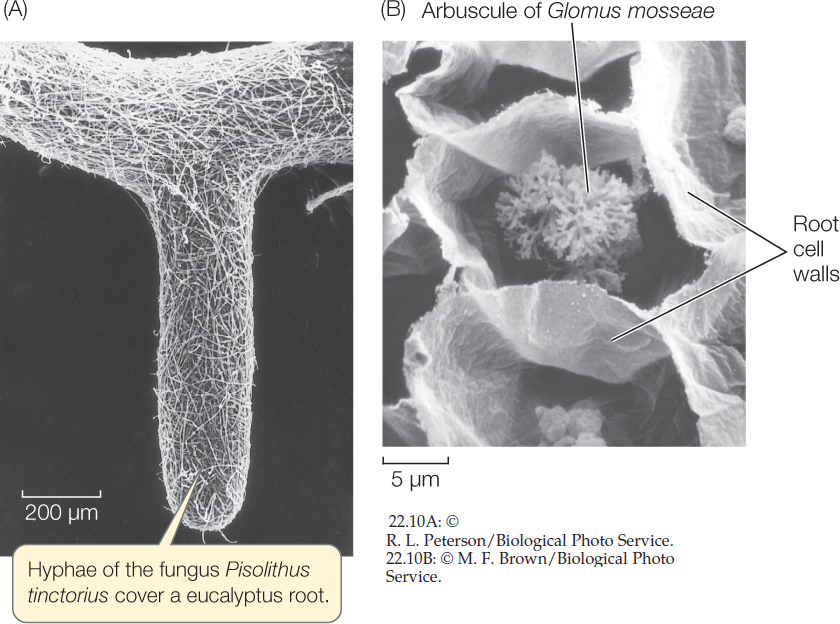
The fungal hyphae of arbuscular mycorrhizae enter the root and penetrate the cell walls of the root cells, forming arbuscular (treelike) structures inside the cell wall but outside the cell membrane. These structures, like the haustoria of parasitic fungi and the contact regions of fungal hyphae and photosynthetic cells in lichens, become the primary site of exchange between plant and fungus (FIGURE 22.10B). As in the ectomycorrhizae, the fungus forms a vast web of hyphae leading from the root surface into the surrounding soil.
LINK
Concept 25.2 contains a detailed description of the role of arbuscular mycorrhizae in plant nutrition
The mycorrhizal association is important to both partners. The fungus obtains needed organic compounds, such as sugars and amino acids, from the plant. In return, the fungus, because of its very high surface area-to-volume ratio and its ability to penetrate the fine structure of the soil, greatly increases the plant’s ability to absorb water and minerals (especially phosphorus). The fungus may also provide the plant with certain growth hormones and may protect it against attack by disease-causing microorganisms.
Plants that have active arbuscular mycorrhizae typically are a deeper green and may resist drought and temperature extremes better than plants of the same species that have little mycorrhizal development. Attempts to introduce some plant species to new areas have failed until a bit of soil from the native area (presumably containing the fungus necessary to establish mycorrhizae) was provided. Trees without ectomycorrhizae do not grow well in the absence of abundant nutrients and water, so the health of our forests depends on the presence of ectomycorrhizal fungi. Many agricultural crops require inoculation of seeds with appropriate mycorrhizal fungi prior to planting. Without these fungi, the plants are unlikely to grow well, or in some cases at all. Certain plants that live in nitrogen-poor habitats, such as cranberry bushes and orchids, invariably have mycorrhizae. Orchid seeds will not germinate in nature unless they are already infected by the fungus that will form their mycorrhizae. Plants that lack chlorophyll always have mycorrhizae, which they often share with the roots of green, photosynthetic plants. In effect, these plants without chlorophyll are feeding on nearby green plants, using the fungus as a bridge.
458
Endophytic fungi protect some plants from pathogens, herbivores, and stress
In a tropical rainforest, 10,000 or more fungal spores may land on a single leaf each day. Some are plant pathogens, some do not affect the plant at all, and some invade the plant in a beneficial way. Fungi that live within aboveground parts of plants without causing obvious deleterious symptoms are called endophytic fungi. Recent research has shown that endophytic fungi are abundant in plants in all terrestrial environments.
Among the grasses, individual plants with endophytic fungi are more resistant to pathogens and to insect and mammalian herbivores than are plants lacking endophytes. The fungi produce alkaloids (nitrogen-containing compounds) that are toxic to animals. The alkaloids do not harm the host plant. In fact, some plants produce alkaloids (such as nicotine) themselves. The fungal alkaloids also increase the ability of grasses to resist stress of various types, including drought (water shortage) and salty soils. Such resistance is useful in agriculture.
The role, if any, of endophytic fungi in most broad-leaved plants is unclear. They may convey protection against pathogens, or they may simply occupy space within leaves without conferring any benefit, but also without doing harm. The benefit, in fact, might be all for the fungus.
CHECKpoint CONCEPT 22.2
- What is the role of fungi in Earth’s carbon cycle?
- Describe the nature and benefits of the lichen association.
- Why do plants grow better when infected with mycorrhizal fungi?
Before molecular techniques clarified the phylogenetic relationships of fungi, one criterion used for assigning fungi to taxonomic groups was the nature of their life cycles—including the types of fruiting bodies they produce. The next section will take a closer look at life cycles in the six major groups of fungi.