Concept 27.1: Most Angiosperms Reproduce Sexually
Most angiosperms (flowering plants) have evolved to reproduce sexually because this strategy results in the genetic diversity that is the raw material for evolution. Sexual reproduction in angiosperms involves mitosis, meiosis, and the alternation of haploid and diploid generations (see Figure 7.3). There are several important differences between sexual reproduction in angiosperms and in vertebrate animals (see Chapter 37):
- Meiosis in plants produces spores, which undergo mitosis to produce gametes; in animals, meiosis usually produces gametes directly.
- In most plants, there are multicellular diploid (sporophyte) and haploid (gametophyte) life stages (alternation of generations); in animals, there is no multicellular haploid stage.
- In plants, the cells that will form gametes (germline cells) develop in the adult organism, usually in response to environmental conditions; in animals, the germline cells are set aside before birth.
LINK
The life cycles of plants have many unique characteristics, which are detailed in Chapter 21
The flower is the reproductive organ of angiosperms
In angiosperms, flowers contain male or female gametophytes or often both. As we saw in Concept 21.5, a generalized flower consists of four concentric groups (whorls) of organs arising from modified leaves: the carpels, stamens, petals, and sepals.
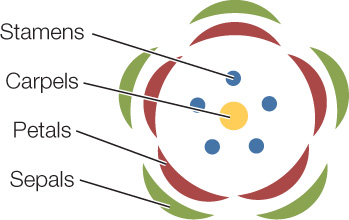
The parts of the flower are all derived from a modified shoot apical meristem.
- Carpels are the female sex organs that contain the developing female gametophytes (inside ovules).
- Stamens are the male sex organs that produce microspores, the first cells of the developing male gametophytes.
The differentiation of these organs is controlled by specific transcription factors (see Figure 14.12).
As you know, flowers come in many shapes, sizes, and forms. Some have both stamens (male sex organs) and carpels (female sex organs); such flowers are termed perfect (FIGURE 27.1A). Imperfect flowers, by contrast, are those with only male or only female sex organs. Male flowers have stamens but not carpels, and female flowers have carpels but not functioning stamens. Some plants, such as corn, bear both male and female flowers on the same individual plant; such species are called monoecious (“one housed”; FIGURE 27.1B). In dioecious (“two-housed”) species, individual plants bear either male-only or female-only flowers; an example is red campion (FIGURE 27.1C).

575
Angiosperms have microscopic gametophytes
FIGURE 27.2 shows the development of angiosperm gametophytes. These haploid, microscopic gamete-producing structures develop from haploid spores in the flower:
- Each female gametophyte (megagametophyte) is called an embryo sac, and it develops inside an ovule. One or more ovules are contained inside an ovary, which is the lower part of a carpel.
- Male gametophytes (microgametophytes), which are also called pollen grains, develop in the anther, which is part of the stamen.

Go to ACTIVITY 27.1 Angiosperm Reproduction
PoL2e.com/ac27.1
Female Gametophyte
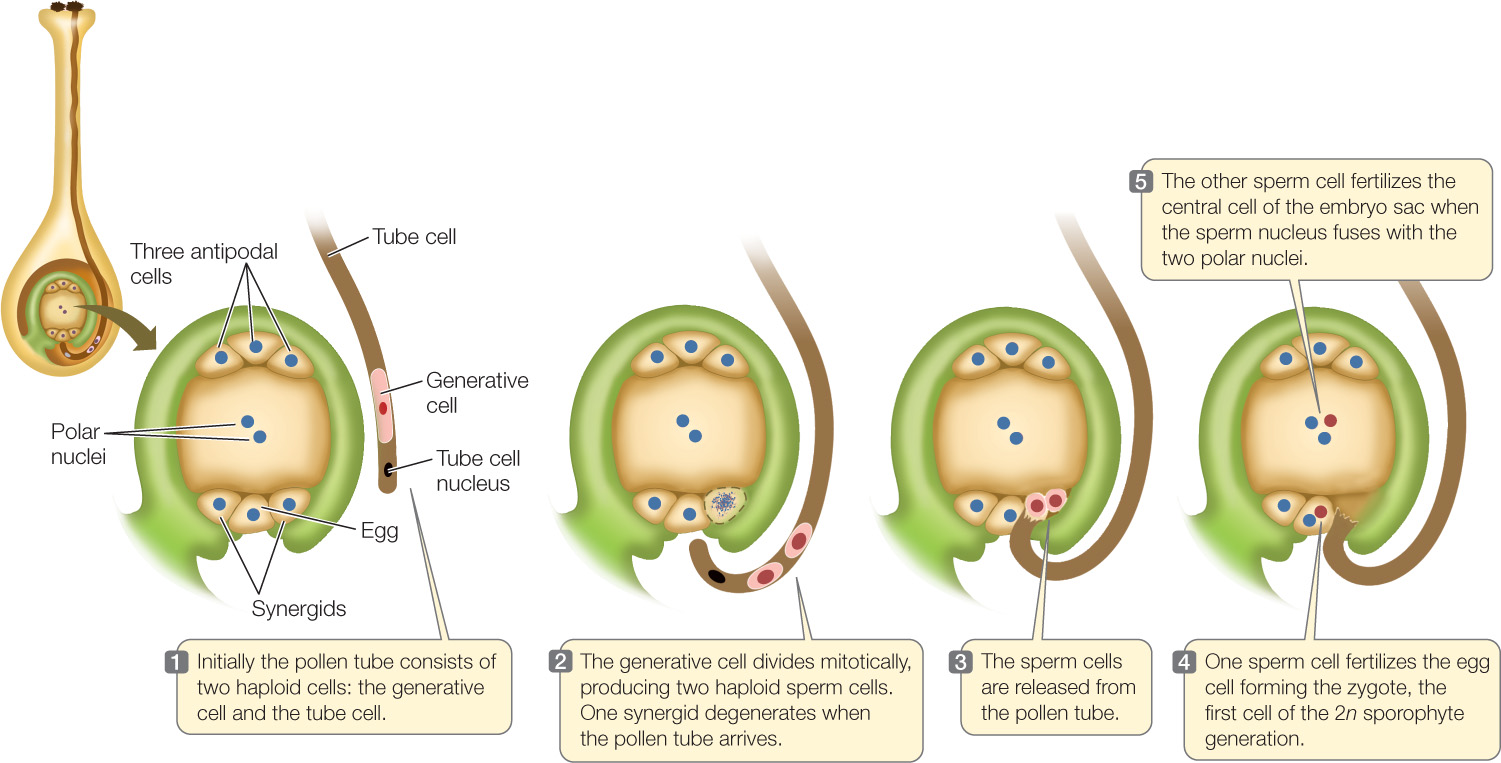
Of the four haploid megaspores resulting from meiosis, three undergo apoptosis (programmed cell death). Typically, the remaining megaspore undergoes three mitotic divisions without cytokinesis, producing eight haploid nuclei, all initially contained within a single cell—three nuclei at one end, three at the other end, and two in the middle. Subsequent cell wall formation leads to an elliptical, seven-celled megagametophyte with a total of eight nuclei:
- At one end of the megagametophyte are three small cells: the egg cell and two cells called synergids. The egg cell is the female gamete, and the synergids participate in fertilization by attracting the pollen tube. The tube enters one of the synergids before the sperm cells are released for fertilization.
- At the opposite end of the megagametophyte are three antipodal cells, which eventually degenerate by apoptosis.
- In the large central cell are two polar nuclei.
The megagametophyte, or embryo sac, is the entire seven-cell, eight-nucleus structure.
Male Gametophyte
The four haploid products of meiosis in the stamen each develop a cell wall and undergo a single mitotic division, producing four two-celled pollen grains (male gametophytes) that are released into the environment. The two cells in a pollen grain have different roles:
- The generative cell divides by mitosis to form two sperm cells that participate in double fertilization. (In some plants this division takes place before pollen is released from the anther, resulting in three-celled pollen grains.)
- The tube cell forms the elongating pollen tube that delivers the sperm to the embryo sac after the pollen grain is transferred to a stigma (part of the female reproductive organ)—a process called pollination.

Go to MEDIA CLIP 27.1 Pollen Germination in Real Time
PoL2e.com/mc27.1
Angiosperms have mechanisms to prevent inbreeding
As we discussed in Concept 21.4, the evolution of pollen in the seed plants made it possible for male gametes to reach the female gametophyte in the absence of liquid water. The transfer of pollen from plant to plant presents its own challenges, however. In the first seed plants to evolve, tiny pollen grains were carried to stigmas by wind. Wind-pollinated flowers have sticky or featherlike stigmas, and they produce a great number of pollen grains because of the low probability of an individual grain encountering a suitable stigma. More recently in evolutionary time, most angiosperms have come to rely on animals such as insects and birds for pollination. Pollen transport by animals greatly increases the probability that pollen will get to a female gametophyte of the same species.
LINK
The evolution of plant–pollinator interactions and their effects on plant speciation are described in Concepts 17.4 and 21.5
Some plants self-pollinate: that is, they are fertilized by pollen that lands on the stigma of a flower on the same plant, or even the same flower. Mendel’s garden peas, for example, reproduce in this way, so he had to remove stamens to control the parentage of his genetic crosses (see Figure 8.1). Self-pollination mitigates the difficulties of wind or animal pollination of a distant flower. However, “selfing” leads to homozygosity, which can reduce the reproductive fitness of offspring (known as inbreeding depression). Most plants have evolved mechanisms that prevent self-pollination and thus self-fertilization.
Separation of Male and Female Gametophytes
Self-fertilization is not possible in dioecious species, which bear only male or female flowers on an individual plant. Pollination in dioecious species is accomplished only when one plant pollinates another. In monoecious plants, which bear both male and female flowers on the same plant, the physical separation of male and female flowers is often sufficient to prevent self-fertilization. Some monoecious species prevent self-fertilization by staggering the development of male and female flowers so they do not bloom at the same time, making these species functionally dioecious.
Genetic Self-Incompatibility
A pollen grain that lands on a stigma of the same plant will germinate and fertilize the female gamete only if the plant is self-compatible (capable of self-pollination). To prevent self-fertilization, many plants are genetically self-incompatible. This mechanism depends on the ability of a plant to determine whether pollen is genetically similar or genetically different from “self.” Rejection of “same-as-self” pollen prevents self-fertilization.
Self-incompatibility in plants is controlled by a cluster of tightly linked genes called the S locus (for self-incompatibility). The S locus encodes proteins in the pollen and style that interact during the recognition process. A self-incompatible species typically has many alleles of the S locus. The pollen phenotype may be determined by its own haploid genotype or by the diploid genotype of its parent plant. In either case, if the pollen expresses an allele that matches one of the alleles of the recipient carpel, the pollen is rejected. Depending on the type of self-incompatibility, the rejected pollen either fails to germinate or the pollen tube is prevented from growing through the style (FIGURE 27.3); either way, self-fertilization is prevented.
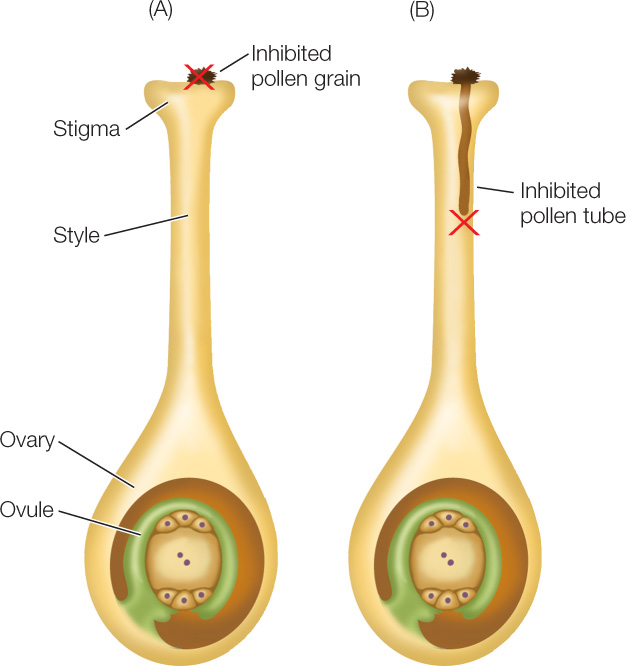
A pollen tube delivers sperm cells to the embryo sac
When a genetically appropriate pollen grain lands on the stigma of a compatible carpel, it germinates. As with seed germination, a key event is water uptake by pollen, in this case from the stigma. Germination involves the development of a pollen tube. The pollen tube either traverses the spongy tissue of the style or, if the style is hollow, grows on the inner surface of the style until it reaches an ovule. The pollen tube typically grows at the rate of 1.5–3 millimeters per hour, taking just 1–2 hours to reach its destination, the female gametophyte.
577
The growth of the pollen tube may be guided in part by a chemical signal produced by the synergids in the female gametophyte. If one synergid is destroyed, the ovule still attracts pollen tubes, but destruction of both synergids renders the ovule unable to attract pollen tubes, and fertilization does not occur. The attractant appears to be species-specific: in some cases, isolated female gametophytes attract only pollen tubes of the same species.
Angiosperms perform double fertilization
The pollen tube grows down the style, under the direction of the haploid tube cell. Meanwhile the generative cell divides once (if it hasn’t already done so) to form two haploid sperm cells. Once the pollen tube reaches the embryo sac, one of the two synergids degenerates, and the two sperm cells are released into its remains. Two fertilization events occur (FIGURE 27.4):
- One sperm cell fertilizes the egg cell, producing the diploid zygote, which forms the new sporophyte embryo.
- The other sperm cell fertilizes the central cell of the embryo sac, when the sperm fuses with the two polar nuclei. This fusion forms a triploid nucleus that undergoes rapid mitosis to form a specialized nutritive tissue, the endosperm. The endosperm’s primary function is nourishment of the developing embryo. In some plants, notably cereals, the endosperm persists in the mature seed and contains reserves that provide chemical building blocks for the germinating seedling while it is underground and cannot perform photosynthesis.

Go to ANIMATED TUTORIAL 27.1 Double Fertilization
PoL2e.com/at27.1
This process of double fertilization is a characteristic feature of angiosperm reproduction. The remaining cells of the female gametophyte—the antipodal cells and the remaining synergid—eventually degenerate, as does the pollen tube nucleus.
Embryos develop within seeds contained in fruits
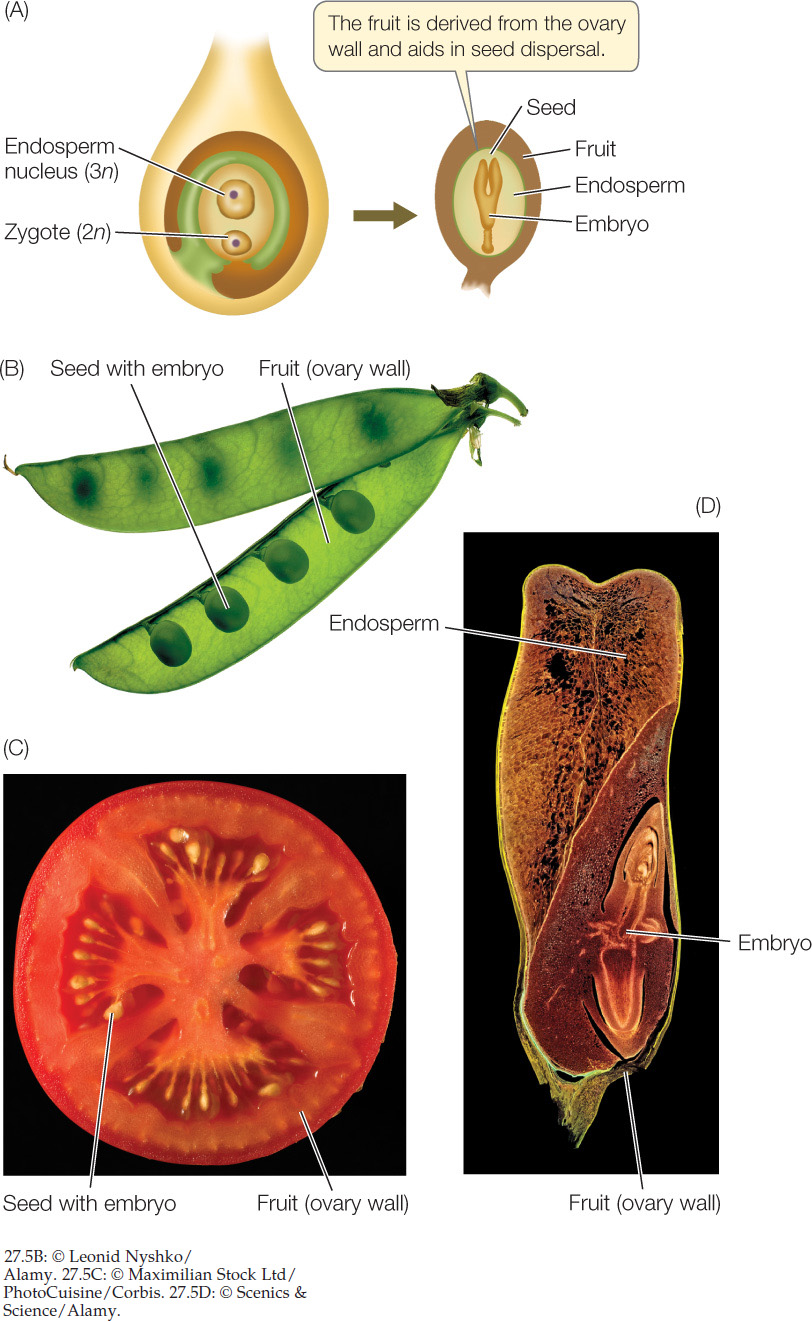
Fertilization initiates the highly coordinated growth and development of the embryo, endosperm, integuments, and carpel. The integuments—tissue layers immediately surrounding the ovule—develop into the seed coat, and the ovary becomes the fruit that encloses the seed (see Figure 27.2).
In Chapter 26 we described the events in plant embryonic development and its hormonal control. As seeds develop, they prepare for dispersal and dormancy by losing up to 95 percent of their water content. You can see this desiccation by comparing corn grains (e.g., popcorn) with ripe corn from the cob or a can. The embryo inside a dry seed contains very little water but is still alive; it has protective proteins that keep its cells in a viscous state.
In angiosperms, the ovary—together with the seeds it contains—develops into a fruit after fertilization has occurred. Fruits have two main functions:
- They protect the seed(s) from damage by animals and infection by microbial pathogens.
- They aid in seed dispersal.
A fruit may consist of only the mature ovary and seeds, or it may include other parts of the flower. Some species produce fleshy, edible fruits, such as peaches and tomatoes, whereas the fruits of other species are dry or inedible (FIGURE 27.5; see also Figure 21.26).
The diverse forms of fruits reflect the varied strategies plants use to disperse their progeny. Because plants cannot move, their progeny need some mechanism for separating themselves from their parent. Fruits are clearly an important way to move seeds away from the parent plant. Wide dispersal of progeny may not always be advantageous, however. If a plant has successfully grown and reproduced, its location is likely to be favorable for the next generation too. Some offspring do indeed stay near the parent, as is the case in many tree species, whose seeds simply fall to the ground. This strategy, however, has several potential disadvantages. If the species is a perennial (see Concept 27.2), offspring that germinate near their parent will be competing with their parent for resources, which may be too limited to support a dense population. Furthermore, even though local conditions were good enough for the parent to produce at least some seeds, there is no guarantee that conditions will still be good the next year, or that they won’t be better elsewhere. Thus in many cases, seed dispersal is vital to a species’ survival.
578
Some fruits help disperse seeds over substantial distances, increasing the probability that at least a few of the many seeds produced by a plant will find suitable conditions for germination and growth to sexual maturity. Some, like those of the thistle, are carried to new locations by wind:

Others attach themselves to animals (or to your clothes and shoes):

Water disperses some fruits; coconuts have been known to float thousands of miles between islands. Seeds swallowed whole by an animal along with fruits such as berries travel through the animal’s digestive tract and are deposited some distance from the parent plant (see Apply the Concept in Chapter 21).
CHECKpoint CONCEPT 27.1
- Make a table that compares the haploid and diploid life stages of a human and a corn plant with regard to (1) the relative sizes of the haploid and diploid stages, (2) how many cells are in the haploid stage, (3) the names of the haploid cells, (4) the time in the life cycle that the haploid stage occurs, (5) the location of the haploid cells, and (6) how the haploid stages develop in males and females. (Refer to Chapter 37 for information on human reproduction.)
- Describe the processes that angiosperms use to ensure a genetically diverse population.
- The two sperm that participate in plant double fertilization are genetically identical. Are the two products of fertilization genetically identical?
579
Now that we have described the cellular aspects of angiosperm reproduction, let’s turn to the events that signal a transition from vegetative growth of the shoot to the formation of flowers.