Concept 27.3: Angiosperms Can Reproduce Asexually
A plant that reproduces asexually produces progeny genetically identical to itself (clones). This mode of reproduction has the disadvantage of not generating the diversity of offspring that sexual reproduction can produce. But if a plant is well adapted to its environment, the parent plant can pass on to all its progeny a combination of alleles that function well in that environment, and which might otherwise be separated by sexual recombination. Furthermore, the plant avoids the costs of producing flowers and the potentially unreliable processes of cross-pollination and seed germination. Many plants are capable of both sexual and asexual reproduction and can switch between the two depending on environmental conditions or the stage in their life cycle.
Angiosperms use many forms of asexual reproduction
Asexual reproduction is often accomplished through the modification of vegetative organs such as roots, stems, or leaves, which is why the term vegetative reproduction is sometimes used to describe asexual reproduction in plants. Another type of asexual reproduction, apomixis, involves flowers but no fertilization.
Vegetative Reproduction
The totipotency of plant cells is dramatically shown when a cell from a fully differentiated organ gives rise to an entire new plant. Strawberries, for example, produce horizontal stems, called stolons or runners, that grow along the soil surface, form roots at intervals, and establish potentially independent plants (see Figure 24.13E). Another example is bamboo, which has underground stems called rhizomes that can form an entire forest (FIGURE 27.12A). Fleshy underground stems, such as potato tubers, produce multiple plants from “eyes” (see Figure 24.13C). Garlic can produce multiple plants from bulbs, which are also modified stems (FIGURE 27.12B). Even leaves can be the source of new plants, as in the succulent Kalanchoe (FIGURE 27.12C).

Plants that reproduce vegetatively often grow in environments such as eroding hillsides, or places where conditions for seed germination are unreliable. Plants with stolons or rhizomes, such as beach grasses, rushes, and sand verbena, are common on coastal sand dunes. Rapid vegetative reproduction enables these plants, once introduced, not only to multiply but also to stay ahead of the shifting sand; in addition, the dunes are stabilized by the extensive network of rhizomes or stolons that develops. Vegetative reproduction is also common in some deserts, where for much of the time the environment is unsuitable for seed germination and the establishment of seedlings.
Vegetative reproduction is efficient in an environment that is stable over the long term. A change in the environment, however, can leave an asexually reproducing species at a disadvantage. For example, the English elm tree (Ulmus procera) reproduces by suckers and is incapable of sexual reproduction. In 1967 a fungal pathogen called Dutch elm disease first struck English elms. The population lacked genetic diversity, and no individuals had genes that would confer resistance to the pathogen. Today the English elm is all but gone from England.
Apomixis
Some plants produce flowers but use them to reproduce asexually rather than sexually. Dandelions, blackberries, some citrus trees, and some other plants reproduce by the asexual production of seeds, called apomixis. In sexual reproduction, seeds form from the union of haploid gametes in the embryo sac. But in apomixis, one of two other things happens:
586
- The cell in the ovule that is supposed to undergo meiosis fails to do so, resulting in a diploid egg cell, which then goes on to form an embryo and seed.
- Diploid cells from the integument surrounding the embryo sac form a diploid embryo sac, and the sac goes on to form an embryo and seed.
In both cases, seed and fruit development proceeds normally. But the genetic consequences are profound: apomixis produces clones.
Some agriculturally important plants are hybrids that are derived from the crossing of two genetically different species. Because these hybrids have two sets of chromosomes that are not homologous to one another, they cannot undergo meiosis and are sterile. This means they cannot form the seeds and fruits that are desired by people by ordinary means. In citrus, apomixis occurs naturally, which gets around this problem. Kentucky blue-grass, a mainstay of lawns, reproduces in this manner as well.
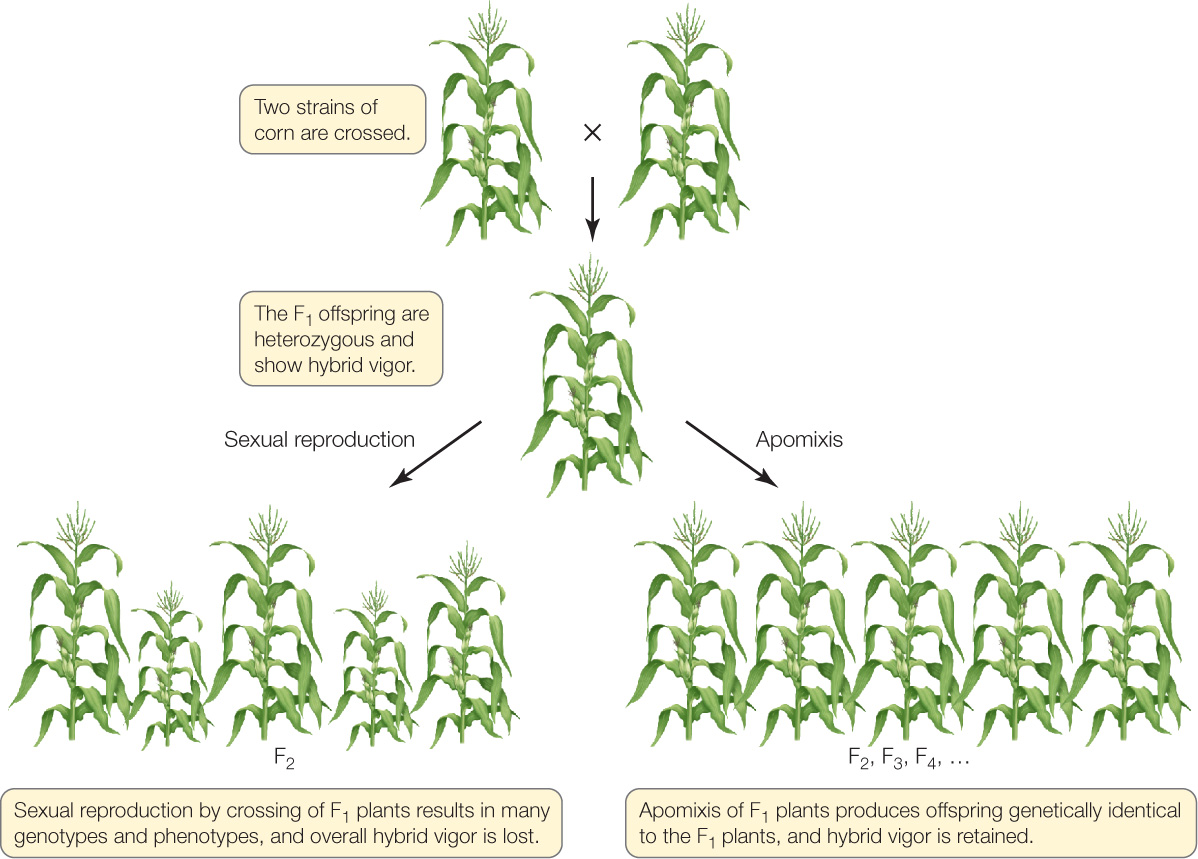
Many important crops, such as corn, are grown as hybrids because the offspring of a cross between two inbred, homozygous genetic strains are often superior to either of their parents, a phenomenon called hybrid vigor (see Concept 8.2). Unfortunately, once farmers have obtained a hybrid with desirable characteristics, they cannot use those plants for further crosses with themselves (selfing) to get more seeds for the next generation. You can imagine the genetic chaos when a hybrid, which is heterozygous at many of its loci (e.g., AaBbCcDdEe, etc.), is crossed with itself: there will be many new combinations of alleles (e.g., AabbCCDdee, etc.), resulting in highly variable progeny. The only efficient and reliable way to reproduce the hybrid is to maintain populations of the original parents to cross again each year. That is exactly what seed companies do. For farmers, it means buying new seeds every year—an expensive proposition.
If a hybrid carried a gene for apomixis, however, it could reproduce asexually, and its offspring would be genetically identical to itself (FIGURE 27.13). New hybrids could be developed that would be adapted to specific environments and could be propagated by the farmers on the spot. Investigators are searching for genes that promote apomixis; these could be introduced into desirable crops and allow them to be propagated indefinitely. Such a gene has been identified in corn, but the yield of the variety that contains it is low.
Vegetative reproduction is important in agriculture
One of the oldest methods of vegetative reproduction used in agriculture consists of simply making cuttings of stems, inserting them in soil, and waiting for them to form roots and become autonomous plants. The cuttings are usually encouraged to root by treatment with auxin, as described in Concept 26.2.
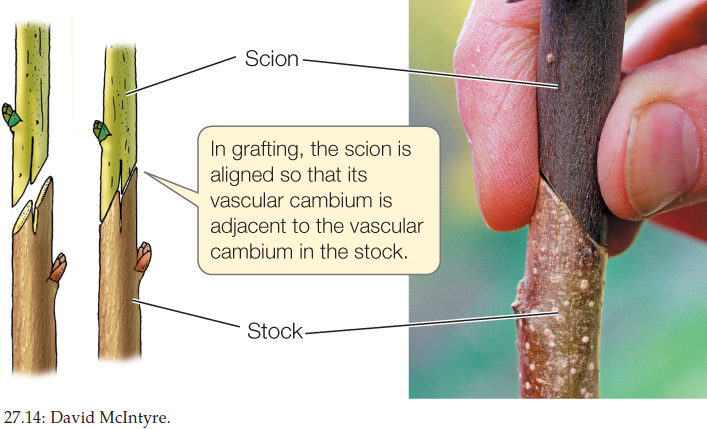
Woody plants can be propagated asexually by grafting: attaching a bud or a piece of stem from one plant to a root or root-bearing stem of another plant. The part of the resulting plant that comes from the root-bearing “host” is called the stock; the part grafted on is the scion (FIGURE 27.14). The vascular cambium of the scion associates with that of the stock, forming a continuous cambium that produces xylem and phloem. The cambium allows the transport of water and minerals to the scion and of photosynthate to the stock. Much of the fruit grown for market in the United States is produced on grafted trees, as are wine grapes. One advantage of this method is that a desirable scion (one that produces good fruit, for example) can be grafted onto a genetically different stock that has other desirable qualities, such as disease-resistant roots.
Another method widely used for asexual plant propagation is meristem culture, in which pieces of shoot apical meristem are cultured on growth media to generate plantlets, which can then be planted in the field. This strategy is vital when uniformity is desired, as in forestry, or when virus-free plants are the goal, as with strawberries and potatoes.
587
CHECKpoint CONCEPT 27.3
- What are the advantages and disadvantages of asexual as compared with sexual reproduction in plants?
- What kinds of gene mutations would you search for that might promote apomixis?
- Most of the navel orange trees that once grew all over southern California are derived by grafting and apomixis from a single tree that still exists in Riverside, CA. How would you prove this?
Question 27.2
How did an understanding of angiosperm reproduction allow floriculturists to develop a commercially successful poinsettia?
ANSWER The poinsettia illustrates well how the concepts of plant physiology and reproduction are used in floriculture. As Albert Ecke found, poinsettias are short-day plants, using the long nights of winter as a signal for flowering (Concept 27.2). In fact, the poinsettia’s inductive dark period of 14 hours was among the first described in a flowering plant. The Eckes’ plants did not flower consistently until they moved their growing operations from urban Los Angeles to an isolated rural region with less light pollution. Today the plants are grown in greenhouses, where the photoperiod is carefully regulated.
A second important factor in the poinsettia’s commercial success was the plant’s growth habit. In Mexico, the wild relatives of cultivated varieties grow up to 3 meters tall, with few branches (FIGURE 27.15; compare these plants with the 0.5 m tall plants in the photograph at the opening of this chapter). Paul Ecke found a variety that was much shorter and formed an attractive, branching plant with axillary shoots. These compact poinsettias were initially propagated asexually by grafting to native plants (Concept 27.3).

The cause of the compact phenotype was assumed to be a dwarfing gene, but several observations cast doubt on this assumption. First, heat treatment of short, branching plants changed their phenotype to the tall, less branched growth habit. Second, heat-treated plants would regain their short, branching growth habit if they were grafted onto untreated short, branching plants. These and other results pointed to a transmissible infective agent as the cause of the short, branching phenotype.
588
Dr. Ing-Ming Lee at the U.S. Department of Agriculture isolated a bacterium called Phytoplasma from the short, branching plants and proved by experiments that it was the causative agent of the compact growth habit. When young plants with the tall growth habit were inoculated with the bacteria, they grew up short and branching. It is suspected that the bacteria change cytokinin levels in the plants, since this hormone regulates branching (see Concept 26.3).
Many new varieties of poinsettias have been generated by conventional sexual reproduction (Concept 27.1). You can look for the phenotypes of these varieties next holiday season. They include stiff stems, new colors (such as white), and longer lifetimes for the colorful bracts.