1.4 EVOLUTION
The themes introduced in the last two sections stress life’s unity: Cells form the basic unit of all life; DNA, RNA, and proteins carry out the molecular functions of all cells; and metabolic reactions build and break down macromolecules. We need only look around us, however, to recognize that for all its unity, life displays a remarkable degree of diversity. We don’t really know how many species share our planet, but reasonable estimates run to 10 million or more. Both the unity and the diversity of life are explained by the process of evolution, change over time.
1.4.1 Variation in populations provides the raw material for evolution, change over time.
Described in detail, evolution by natural selection calls on complex mathematical formulations, but at heart its main principles are simple, indeed unavoidable. When there is variation within a population of organisms, and when that variation can be inherited (that is, when it can be passed from one generation to the next), the variants best suited for growth and reproduction in a given environment will contribute disproportionately to the next generation. As Darwin recognized, farmers have used this principle for thousands of years to select for crops with high yield or improved resistance to drought and disease. It is how people around the world have developed breeds of dog ranging from terriers to huskies (Fig. 1.15). And it is why antibiotic resistance is on the rise in many disease-causing microorganisms. Life has been shaped by evolution since its origin, and the capacity for Darwinian evolution may be life’s most fundamental property.

1-13
The apples in the bin from which you made your choice didn’t all look alike. Had you picked your apple in an orchard, you would have seen that different apples on the same tree looked different—some smaller, some greener, some misshapen, a few damaged by worms. Such variation is so commonplace that we scarcely pay attention to it. Variation is observed among individuals in virtually every species of organism. Variation that can be inherited provides the raw material on which evolution acts.
The causes of variation among individuals within a species are usually grouped into two broad categories. Variation among individuals is sometimes due to differences in the environment; this is called environmental variation. Among apples on the same tree, some may have good exposure to sunlight, some may be hidden in the shade, some were lucky enough to escape the female codling moth, whose egg develops into a caterpillar that eats its way into the fruit. These are all examples of environmental variation.
The other main cause of variation among individuals is differences in the genes that are transmitted from parents to their offspring; this is known as genetic variation. Genes are composed of DNA and contain coded information that instructs the cell to produce specific types of RNA and protein. Differences among individuals’ DNA lead to differences among the individuals’ RNA and proteins, which affect the molecular functions of the cell and ultimately can lead to physical differences that we can observe. Genetic differences among apples produce varieties whose mature fruits differ in taste and color, such as the green Granny Smith, the yellow Golden Delicious, and the scarlet Red Delicious.
But even on a single tree, each apple contains seeds that are genetically distinct, because the apple tree is a sexual organism. Bees carry pollen from the flowers of one tree and deposit it in the flowers of another, enabling the sperm inside pollen grains to fertilize egg cells within that single flower. All the seeds on an apple tree contain shared genes from one parent, the tree on which they developed. But they contain distinct sets of genes contributed by sperm transported in pollen from other trees. In all sexual organisms, fertilization produces unique combinations of genes, which explains in part why sisters and brothers with the same parents can be so different from one another.
Genetic variation arises ultimately from mutations. A few mutations arise in each generation as a result of errors in DNA replication and environmental factors such as UV radiation. Most genetic variation, however, is due to mutations that occurred in previous generations. To put a human face on this, consider that lung cancer can reflect either environmental insult from cigarette smoking, for example, or an inherited genetic susceptibility.
In nature, most mutations that are harmful to growth and reproduction die out after a handful of generations. Those that are neither harmful nor beneficial can persist for hundreds or thousands of generations. And those that are beneficial to growth and reproduction can gradually become incorporated into the genetic makeup of every individual in the species. That is how evolution works: The genetic makeup of a population changes over time.
1.4.2 Evolution predicts a nested pattern of relatedness among species, depicted as a tree.
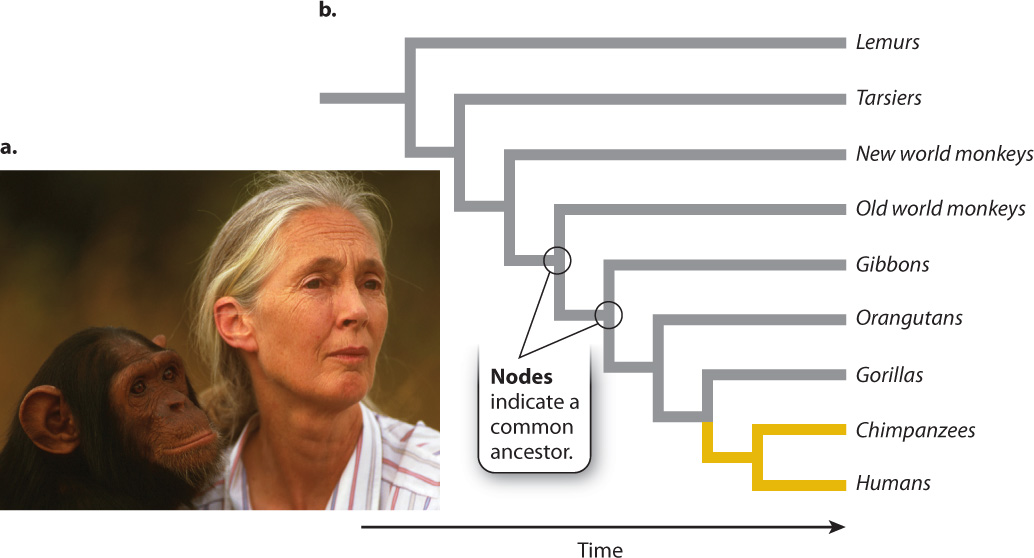
Evolutionary theory predicts that new species arise by the divergence of populations through time from a common ancestor. As a result, closely related species are likely to resemble each other more closely than they do more distantly related species. You know this to be true from common experience. All of us recognize the similarity between a chimpanzee’s body and our own (Fig. 1.16a), and biologists have long known that we share more features with chimpanzees than we do with any other species.
1-14
Humans and chimpanzees, in turn, share more features with gorillas than they do with any other species. And humans, chimpanzees, and gorillas share more features with orangutans than they do with any other species. And so on. We can continue to include a widening diversity of species, successively adding monkeys, lemurs, and other primates, to construct a set of evolutionary relationships that can be depicted as a tree (Fig. 1.16b). In this tree, the tips or branches on the right represent different groups of organisms, nodes (where lines split) represent the most recent common ancestor, and time runs from left to right.
Evolutionary theory predicts that primates show a nested pattern of similarity, and this is what morphological and molecular observations reveal. We can continue to add other mammals, and then other vertebrate animals, to our comparison, in the process generating a pattern of evolutionary relationships that forms a larger tree, with the primates confined to one limb. And using molecular comparisons among species, we can generate still larger trees, ones that include plants as well as animals and the full diversity of microscopic organisms. Biologists call the full set of evolutionary relationships among all organisms the tree of life. This tree, illustrated in Fig. 1.17, has three major branches representing the three domains mentioned earlier and is made up mostly of microorganisms. The last common ancestor of all living organisms, which form a root to the tree, is thought to lie between the branch leading to Bacteria and the branch leading to Archaea and Eukarya. The plants and animals so conspicuous in our daily existence make up only two branches on the eukaryotic limb of the tree.
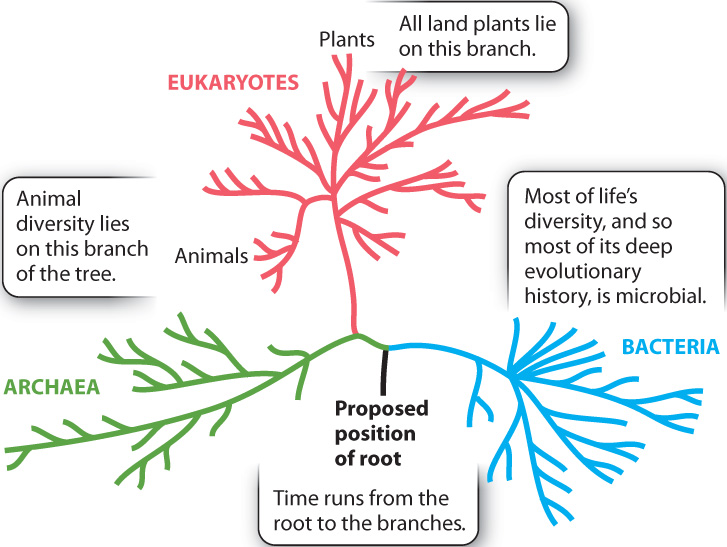
The tree of life makes predictions for the order of appearance of different life-forms in the fossil record. For example, primates should show up before humans, and the first mammals should appear even earlier. The tree also predicts that all records of animal life should be preceded by a long interval of microbial evolution. As we will see in subsequent chapters, these predictions are met by the geologic record.
Shared features, then, sometimes imply inheritance from a common ancestor. In practice, molecular comparisons and fossils show that the close similarity between humans and chimpanzees reflects descent from a common ancestor that lived about 6 million years ago. Their differences reflect what Darwin called “descent with modification”—evolutionary changes that have accumulated over time since the two lineages split. For example, the flat face of humans, our small teeth, and our upright posture all evolved within our ancestors after they diverged from the ancestors of chimpanzees. As discussed in Chapter 24, fossils document the accumulation of uniquely human features over the past 6 million years. At a broader scale, the fundamental features of molecular function and cell organization that are shared by all organisms reflect inheritance from a common ancestor that lived billions of years ago. And the nested patterns of similarity and differences that characterize the many branches on the tree of life have formed through the continuing action of evolution since the time of our earliest ancestors.
1-15
Nearly four decades ago, the geneticist Theodosius Dobzhansky wrote, “Nothing in biology makes sense except in the light of evolution.” For this reason, evolution permeates discussions throughout this book, whether we are explaining the molecular biology of cells, how organisms function and reproduce, how species interact in nature, or the remarkable biological diversity of our planet.
1.4.3 Evolution can be studied by means of experiments.
Both the patterns of similarity among living organisms and the succession of fossils in the geologic record fit the predictions of evolutionary theory. Can we actually capture evolutionary processes in action? One way to accomplish this is in the laboratory. Bacteria are ideal for these experiments because they reproduce rapidly and can form laboratory populations with millions of individuals. Large population size means that mutations will form in nearly every generation, even though the probability that any individual cell will acquire a mutation is small. (In contrast to bacteria, think about trying evolutionary experiments on elephants!)
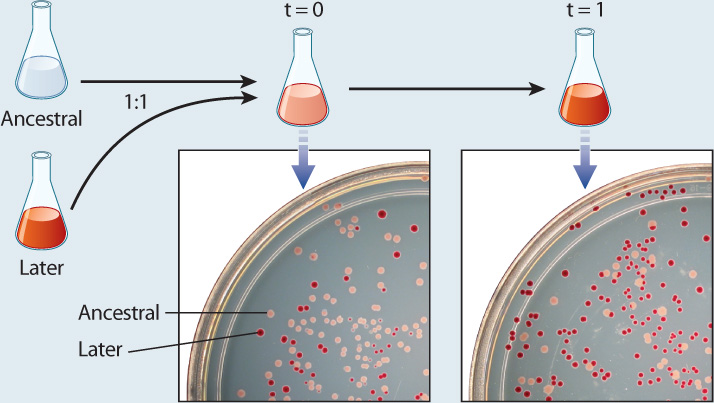
One such experiment is illustrated in Fig. 1.18. The microbiologists Santiago Elena and Richard Lenski grew populations of the common intestinal bacterium Escherichia coli in a liquid medium, with the organic acid succinate as the only source of food. In general, E. coli cells can feed on succinate poorly if at all, leading the researchers to hypothesize that any bacterium with a mutation that increased its ability to utilize succinate would reproduce at a faster rate than other bacteria in the population. The key questions were whether bacteria from later generations differed from those of earlier generations—that is, did evolution occur in the population?—and in particular whether the bacteria had evolved an improved ability to metabolize succinate.
FIG. 1.18: Can evolution be demonstrated in the laboratory?
BACKGROUND Escherichia coli is an intestinal bacterium commonly used in the laboratory. It grows poorly in liquid media where the only source of food is succinate. Santiago Elena and Richard Lenski wondered whether E. coli grown for 20,000 generations in succinate would evolve in ways that improved their ability to metabolize this compound.
HYPOTHESIS Any bacterium with a random mutation that happens to increase its ability to utilize succinate will reproduce at a faster rate than other bacteria in the population. Over time, such a mutant will increase in frequency relative to other types of bacteria, thereby demonstrating evolution in a bacterial population.
EXPERIMENT Cells of E. coli can be frozen in liquid nitrogen, which keeps them in a sort of suspended animation in which no biological processes take place, but the cells survive. At the beginning of the experiment, the researchers froze a large number of samples of the starting bacteria (“ancestral”). As the experiment progressed, they took samples of the bacterial populations at intervals (“later”) and grew them together with a thawed sample of the starting bacteria in succinate. They then compared the rate of growth of the ancestral bacteria to that of the bacteria taken at later time points.
RESULTS At each time interval, the cells from later time points grew more rapidly than the ancestral cells when the two populations were grown together in succinate.
Over time, the bacteria grown in succinate showed more and more improvement in growth compared to the starting bacteria.
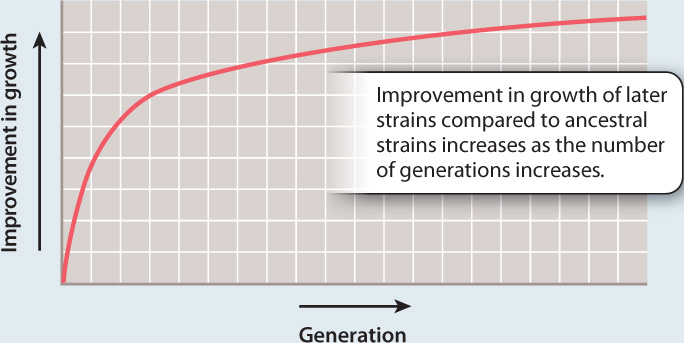
CONCLUSION Evolution occurred in the population: the bacteria evolved an improved ability to metabolize succinate.
FOLLOW-UP WORK Other experimental studies have shown that bacteria are able to evolve adaptations to a wide variety of environmental conditions.
SOURCE Elena, S. F., and R. E. Lenski. 2003. “Evolution Experiments with Microorganisms.” Nature Review Genetics 4:457–469.
1-16
In fact, the bacteria did evolve an improved ability to use succinate, which is demonstrated by the results shown in Fig. 1.18. This experiment illustrates several key points. It shows how experiments can be used to test hypotheses and, of course, it shows evolution in action. Not only did the experiments demonstrate evolutionary adaptation of E. coli populations to a specific laboratory environment, but follow-up studies of their DNA identified differences in genetic makeup that resulted in the observed changes. Many experiments of this general type have been carried out, applying the scientific method to demonstrate how bacteria adapt through mutation to any number of imposed environments.
Experiments in laboratory evolution help us to understand how life works, and they have an immensely important practical side as well. They allow biologists to develop new and beneficial strains of microorganisms that, for example, remove toxins from lakes and rivers and show how some of our worst pathogens develop resistance to drugs designed to eliminate them.