9.2 TYPES OF CELL SIGNALING
In prokaryotes and unicellular eukaryotes, cell communication is communication between individuals. In complex multicellular eukaryotes, cell signaling involves communication between cells that make up an individual. The same principles apply in both instances, but there are differences to be considered. In multicellular organisms, the distance between communicating cells varies considerably. In addition, many cells in multicellular organisms are physically attached to one another, in which case the signaling molecule is not released from the signaling cell. In this section, we explore communication over long and short distances, as well as communication among cells that are physically associated with one another.
9.2.1 Endocrine signaling acts over long distances.
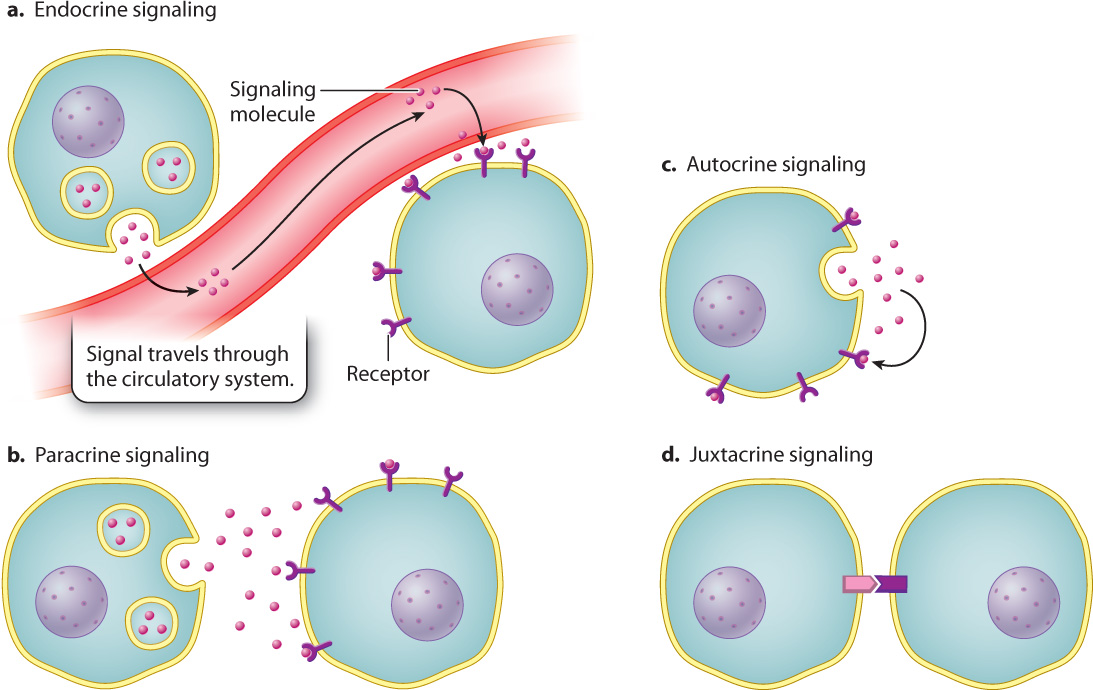
Signaling molecules released by a cell may have to travel great distances in the body, usually via the circulatory system, to reach their target responding cells. Signaling by means of molecules that travel through the bloodstream is called endocrine signaling (Fig. 9.4a and Chapter 38).
9-4
Familiar examples of endocrine-signaling molecules include the mammalian steroid hormones estradiol (an estrogen) and testosterone (an androgen). These hormones travel from the ovaries and the testes, respectively (although there are other minor sources of these hormones), through the bloodstream, to target cells in various tissues throughout the body. The increased amount of estrogen in girls during puberty causes the development of breast tissue and the beginning of menstrual cycles. The increased amount of testosterone in boys during puberty causes the growth of muscle cells, the deepening of the voice, and the growth of facial hair (Chapter 42).
Question Quick Check 1
1W0aKyXZ93L7nOrv7OTbw9rMIX9JwBivIkSb/HAihOIEt3xIGXEUxm2nYSvoUNQ/8Sv8BGwKsXR9UyZ08p+D1PevYiiXAHpfL6VDv29p4Cad3LsdYUak0ADUxpp2okdwXJOovnkq18H256G1LaDVh5SJmjZbkYYWScoJ2tcQW9cdPtoi10RJjyYOorbApaUFcfOWmznRQc/fFfMF0aeDtRZ1/Mo=9.2.2 Paracrine and autocrine signaling act over short distances.
In contrast to endocrine signaling, paracrine signaling (Fig. 9.4b) involves two cells that are close together. In this case, a signaling molecule needs to diffuse only a short distance to the nearest neighboring cell in order to bind its receptor and deliver its message. Signaling molecules in paracrine communication travel distances of up to 20 cell diameters, or a few hundred micrometers. In some cases, signaling molecules may be secreted by a cell and then bind to receptors on the very same cell, as we saw in the case of pneumococcal cells. Such cases, where signaling cell and responding cell are one and the same, are examples of autocrine signaling (Fig. 9.4c). Paracrine and autocrine signaling are especially important to multicellular organisms during the development of the embryo (Chapter 20).
In paracrine signaling, the signal is usually a small, soluble molecule, such as a growth factor. The first growth factor discovered was found by scientists who were attempting to understand how to maintain cultures of cells in the laboratory for use in their research. For decades, medical research has worked with cells maintained in culture. Initially, these cultured cells had limited use because they failed to divide outside the body unless they were supplied with unidentified factors from mammalian blood serum. In 1974, American scientists Nancy Kohler and Allan Lipton discovered that these factors are secreted by platelets (Fig. 9.5). Consequently, the first growth factor was named “platelet-derived growth factor,” or PDGF. We now know of scores of proteins secreted by cells that function as growth factors, and in most cases their effects are confined to neighboring cells. These effects include the stimulation of cell growth, cell division, and changes in gene expression.
FIG. 9.5: Where do growth factors come from?
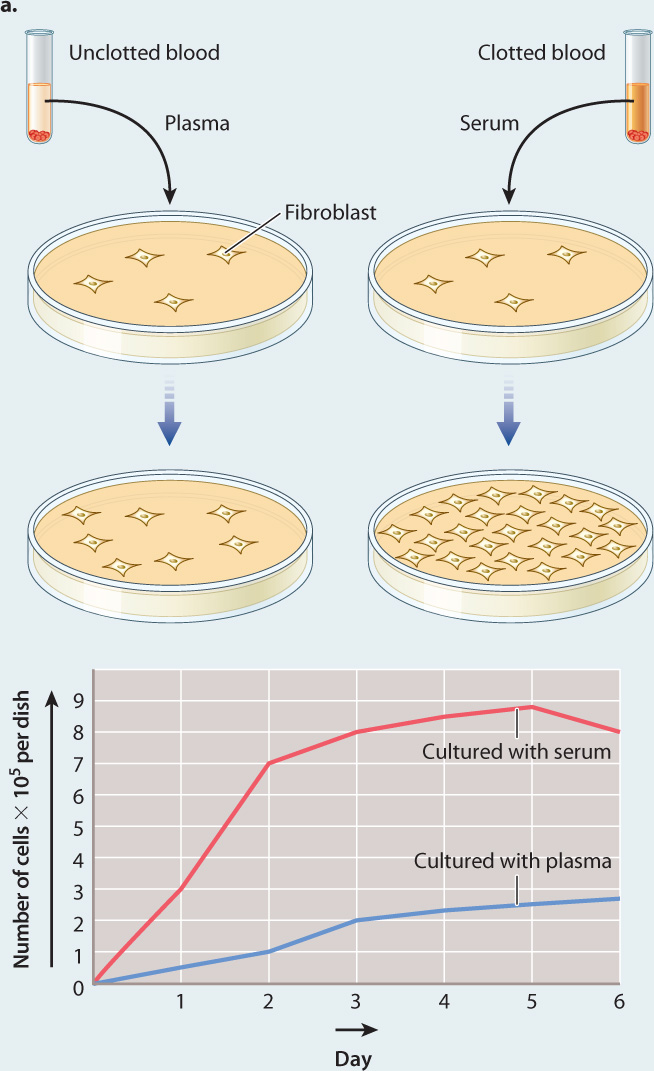
BACKGROUND Cells can be grown outside the body in culture. However, they survive and grow well only under certain conditions. Researchers hypothesized that there are substances that are required for growth of cells in culture, but the nature and source of these substances was unknown. A key insight came from the observation that chicken cells grew much better if they were cultured in the presence of blood serum rather than blood plasma. Blood serum is the liquid component of blood that is collected after blood has been allowed to clot. Blood plasma is also the liquid component of blood, but it is collected from blood that has not clotted. American biologists Nancy Kohler and Allan Lipton were interested in identifying the source of the factor in blood serum that allows cells to survive in culture.
HYPOTHESIS Since Kohler and Lipton knew that clotting depends on the release of substances from platelets, they hypothesized that a growth-promoting factor was activated or introduced into the blood by platelets during the clotting reaction.
EXPERIMENT 1 The investigators first confirmed earlier observations using cells called fibroblasts, which they took from mice. They cultured two sets of fibroblasts in small plastic dishes. To one of the cultures they added serum; to the other culture they added plasma. Then they monitored the rate of cell division in both culture dishes for nine days.
RESULTS 1 They observed that the rate of cell division in the fibroblasts cultured in serum was far greater than that of the cells cultured in plasma, as expected (Fig. 9.5a).
EXPERIMENT 2 To see if the factor is released directly from platelets, they prepared a solution of proteins made from purified platelets, added these proteins to cultured fibroblasts, and measured cell growth.
RESULTS 2 They found that the solution of platelet proteins also caused an increase in the growth of fibroblasts compared to the growth of fibroblasts in plasma (Fig. 9.5b).
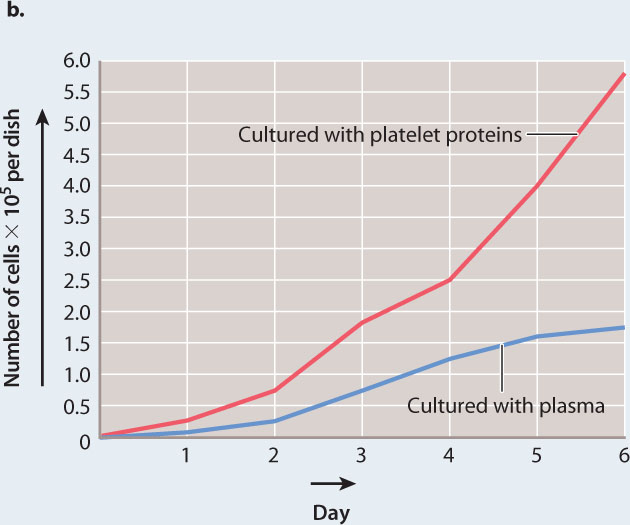
CONCLUSION Kohler and Lipton concluded that the growth-promoting factor is a protein that is released by platelets on activation and clot formation, and therefore normally present in serum.
FOLLOW-UP WORK Subsequent work over the next few years by these and other scientists resulted in the purification and characterization of the growth factor we know today as platelet-derived growth factor, or PDGF.
SOURCE Kohler, N., and A. Lipton. 1974. “Platelets as a Source of Fibroblast Growth-Promoting Activity.” Experimental Cell Research 87:297–301.
Growth factors secreted by cells in an embryo work over short distances to influence the kind of cells their neighbors will become. In this way, they help shape the structure of the adult’s tissues, organs, and limbs. For example, in developing vertebrates, paracrine signaling by the growth factor Sonic Hedgehog (yes, it’s named after a video game character) ensures that the motor neurons in your spinal cord are located properly, that the bones of your vertebral column form correctly, and that your thumb and pinky fingers are on the correct sides of your hands.
A specialized form of paracrine signaling in many organisms is the communication between neurons (nerve cells), or between neurons and muscle cells (Chapter 35). When neurotransmitters, a type of signaling molecule, are released from a neuron, they diffuse across a very small space, called a synapse, between the signaling cell and the responding cell. If the adjacent cell is a neuron, it often responds by transmitting the nerve impulse further and then releasing additional neurotransmitters. If the responding cell is a muscle cell, it often responds by contracting.
9-5
9.2.3 Juxtacrine signaling depends on direct cell–cell contact.
In some cases, a cell communicates with another cell through direct physical contact, without a chemical signal that diffuses or circulates through an external medium. This type of communication is called contact-dependent signaling, or juxtacrine signaling (Fig. 9.4d). It requires that the two communicating cells be in physical contact with each other. A transmembrane protein on the surface of one cell acts as the signaling molecule, and a transmembrane protein on the surface of an adjacent cell acts as the receptor.
As an example of juxtacrine signaling, let’s look at the development of the central nervous system of vertebrate animals. In the brain and spinal cord, neurons transmit information in the form of electrical signals from one part of the body to another. The neurons in the central nervous system are greatly outnumbered by supporting cells, called glial cells, that nourish and insulate the neurons. Both the neurons and the glial cells start out as similar cells in the embryo, but some of these undifferentiated cells become neurons and many more become glial cells.
9-6
For reasons that are not entirely clear, the amount of a transmembrane protein called Delta dramatically increases on the surface of some of these undifferentiated cells. Delta binds to a transmembrane protein called Notch on the surface of adjacent cells. In this case, the signaling cell is the cell with elevated levels of Delta protein. The Delta protein in turn is the signaling molecule, and Notch is its receptor. The signal Delta sends to the adjacent, undifferentiated cell through Notch is a message to become a glial cell and not a neuron. Because one signaling cell sends this same message to all the cells it touches, it is easy to understand how there can be so many more glial cells than neurons in the central nervous system.
As you can see from these examples, the ways in which cells of a developing embryo communicate with one another and the way neurons signal muscle cells or other neurons are based on the same principles at work in DNA uptake by pneumococcal cells in response to a peptide signal. All these communications are based on small-molecule signals that are exchanged between cells. These signaling molecules are the language of cellular communication.