9.3 RECEPTORS AND RECEPTOR ACTIVATION
Receptors are proteins that receive and interpret information carried by signaling molecules. Regardless of the distance between communicating cells, a message is received by a cell when the signaling molecule binds noncovalently to its appropriate and usually highly specific receptor protein. For this reason, the signaling molecule is often referred to as a ligand (from the Latin ligare, which means “to bind”). The signaling molecule binds to a specific location on the receptor protein called the ligand-binding site.
Almost without exception, the binding of a signaling molecule to the ligand-binding site of a receptor causes a conformational change in the receptor. We say that the conformational change “activates” the receptor because it is through this change that the receptor passes the message from the signaling molecule to the interior of the cell. In many ways, this change in receptor shape is similar to the change that occurs when a substrate binds to the active site of an enzyme (Chapter 6). The conformational change in the receptor ultimately triggers various chemical reactions or other changes in the cytosol, and is therefore a crucial step in the reception and interpretation of communications from other cells.
9.3.1 Receptors can be on the cell surface or in the interior of the cell.
The location of a particular receptor in a cell depends largely on whether the signaling molecule is polar or nonpolar (Fig. 9.6). Many signaling molecules, such as the growth factors we just encountered, are small, polar proteins that cannot pass through the hydrophobic core of the plasma membrane. The receptor proteins for these signals are on the outside surface of the responding cell (Fig. 9.6a).
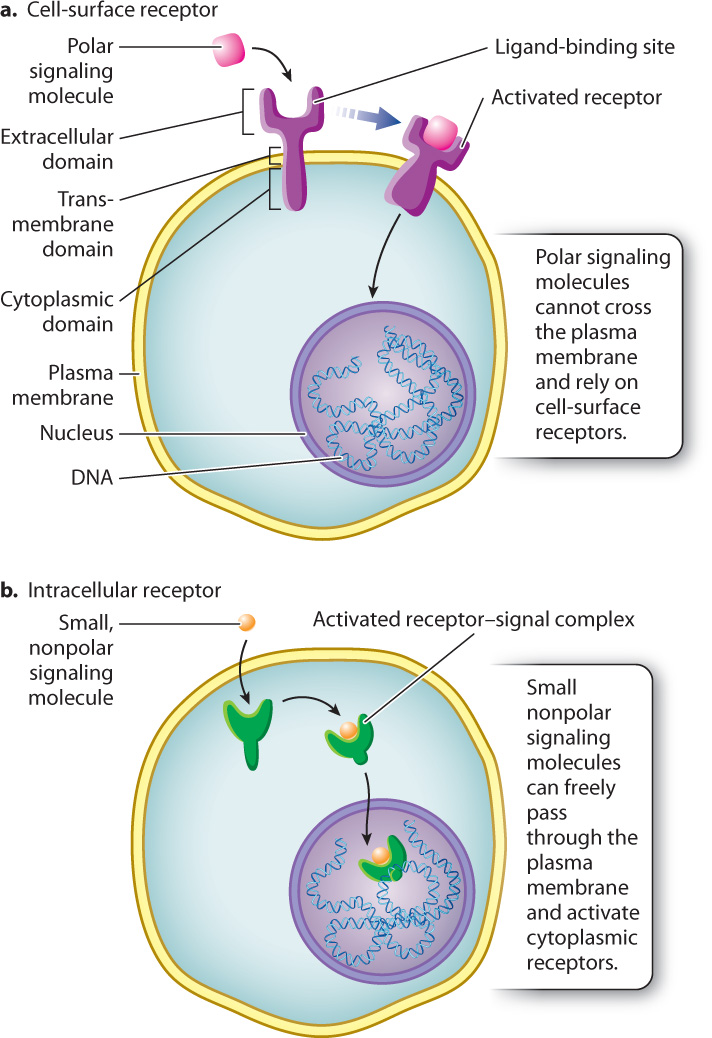
Receptor proteins for growth factors and other polar ligands are transmembrane proteins with an extracellular domain, a short transmembrane domain, and a cytoplasmic domain. When a signaling molecule binds to the ligand-binding site in the extracellular domain, the entire molecule, including the cytoplasmic domain of the receptor, undergoes a conformational change, and as a result the molecule is activated. In this way, the receptor acts as a bridge between the inside and outside of the responding cell that carries the message of the hydrophilic signal across the hydrophobic core of the plasma membrane. The type of receptor determines which signaling pathway is activated and ultimately determines how the cell responds to the signaling molecule. We discuss several examples of signaling pathways in section 9.4.
9-7
Some nonpolar signaling molecules, such as the steroid hormones involved in endocrine signaling, don’t need a receptor on the cell surface in order to relay information to the interior of the cell. Since steroids are derived from cholesterol, they are hydrophobic and pass easily through the hydrophobic core of the phospholipid bilayer and into the target cell. Once inside, steroid hormones bind to receptor proteins located in the cytosol or in the nucleus to form receptor–steroid complexes (Fig. 9.6b). Steroid–receptor complexes formed in the cytosol enter the nucleus, where they act as transcriptional regulators to control the expression of specific genes. Steroid receptors located in the nucleus are often already bound to DNA and need only to bind their steroid counterpart to become active.
The importance of steroid hormones and their receptors cannot be overstated. Sex hormones, glucocorticoids (which raise blood glucose levels), and ecdysone (involved in insect molting) are examples of steroid hormones. However, since the majority of information received by cells is transmitted across the plasma membrane through transmembrane receptors, we focus our attention on the sequence of events that takes place when receptors on the surface of cells bind their ligands.
9.3.2 There are three major types of cell-surface receptor, which act like molecular switches.
As we saw earlier, receptor activation occurs after a signaling molecule binds to the ligand-binding site on its receptor. Many receptors, in fact, act as binary molecular switches, existing in only two alternative states, either “on” or “off,” like a light switch without a dimmer (Fig. 9.7a). Receptors behave similarly to a light switch. When bound to their signaling molecule, the molecular switch is turned on. When the signaling molecule is no longer bound, the switch is turned off.
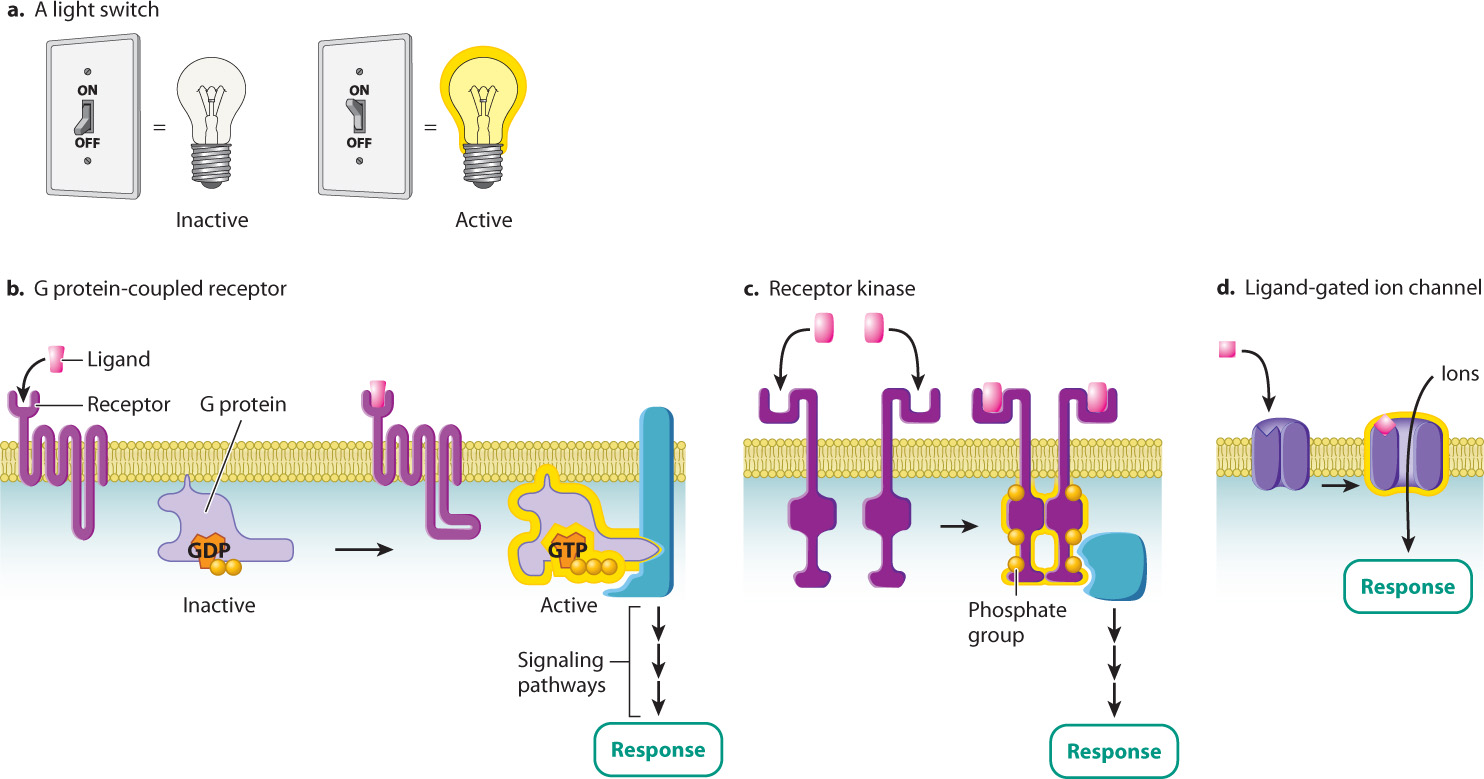
There are thousands of different receptor proteins on the surface of any given cell. Most of them can be placed into one of three groups according to the way they are activated. The first type of cell-surface receptor is called a G protein-coupled receptor (Fig. 9.7b). As its name suggests, this type of receptor couples to, or associates with, G proteins, proteins that bind to the guanine nucleotides GTP and GDP. When a G protein is bound to GTP, it is active, and when it is bound to GDP, it is inactive. When a ligand binds to a G protein-coupled receptor, the receptor binds to and activates the G protein by replacing GDP with GTP. As long as the G protein is bound to GTP, it is in the “on” position and the signal is propagated.
Question Quick Check 2
5xHiiCgMjw7GBb9NR0u5SrjzPuAnOymm37T7200RGMaamt49wKin2Q2lAtp4k1kwcoJrvM6zkwS8YDZuCoTyu6i8dBCDtx325ysF6ByvNpbh+6gw2MoZvQ==The second type of receptor is an enzyme called a receptor kinase (Fig. 9.7c). A kinase is an enzyme that adds a phosphate group to another molecule in a process called phosphorylation. Phosphatases have the opposite effect and remove a phosphate group, a process called dephosphorylation. Usually (but not always), when a protein is phosphorylated by a kinase, it becomes active and is switched on. When the protein is dephosphorylated by a phosphatase, it is switched off and becomes inactive.
Receptors in the third group, the ligand-gated ion channels, alter the flow of ions across the plasma membrane when bound by their ligand (Fig. 9.7d). Recall from Chapter 5 that channel proteins help ions and other molecules diffuse into and out of the cell by providing a hydrophilic pathway through the hydrophobic core of the phospholipid bilayer. Most of the time, the channels are closed to prevent the free movement of ions across the membrane. However, when a signaling molecule binds to the extracellular ligand-binding site of the channel protein, the channel undergoes a conformational change that opens it and allows ions to flow in and out. The channel remains open as long as the signaling molecule remains bound. This type of signaling is especially important for neurons and muscle cells, since their primary functions depend on a rapid change in ion transport across the plasma membrane.