11.1 CELL DIVISION
Cell division is the process by which a single cell becomes two daughter cells. While this process may seem simple, successful cell division must satisfy several important requirements. First, the two daughter cells must each receive the full complement of genetic material (DNA) present in the single parent cell. Second, the parent cell must be large enough to divide in two and still contribute sufficient cytoplasmic components such as proteins, lipids, and other macromolecules to each daughter cell. Satisfying these requirements means that key cellular components must be duplicated before cell division takes place. This duplication of material is achieved in a series of steps that constitute the life cycle of every cell. When you think of a life cycle, you might think of various stages beginning with birth and ending with death. In the case of a single cell, the life cycle begins and ends with cell division.
In this section, we explore the different mechanisms by which prokaryotic and eukaryotic cells divide. Prokaryotic cells divide by binary fission. When eukaryotic cells divide, they first divide the nucleus during mitosis, then divide the cytoplasm into two daughter cells during cytokinesis. As we discuss, it is likely that mitosis evolved from binary fission.
11.1.1 Prokaryotic cells reproduce by binary fission.
The majority of prokaryotic cells, namely bacteria and archaeons, divide by binary fission. In this form of cell division, a cell replicates its DNA, increases in size, and divides into two daughter cells. Each daughter cell receives one copy of the replicated parental DNA. The molecular mechanisms that drive binary fission have been studied most extensively in bacteria. The process of binary fission is similar in archaeons, as well as in chloroplasts and mitochondria, organelles within plant and animal cells which evolved from free-living prokaryotic cells (Chapters 5 and 27).
Let’s consider the process of binary fission in the intestinal bacterium Escherichia coli (Fig. 11.1). The circular genome of E. coli is attached by proteins to the inside of the plasma membrane. DNA replication is initiated at a specific location on the circular DNA molecule and proceeds in opposite directions around the circle. The result is two DNA molecules, both attached to the plasma membrane. The cell then grows. As the cell elongates, the two DNA attachment sites move apart. When the cell is about twice its original size and the DNA molecules are well separated, a constriction forms at the midpoint of the cell. Eventually, new membrane and cell wall are synthesized at the site of the constriction, dividing the single cell into two. The result is two daughter cells, each having the same genetic material as the parent cell.
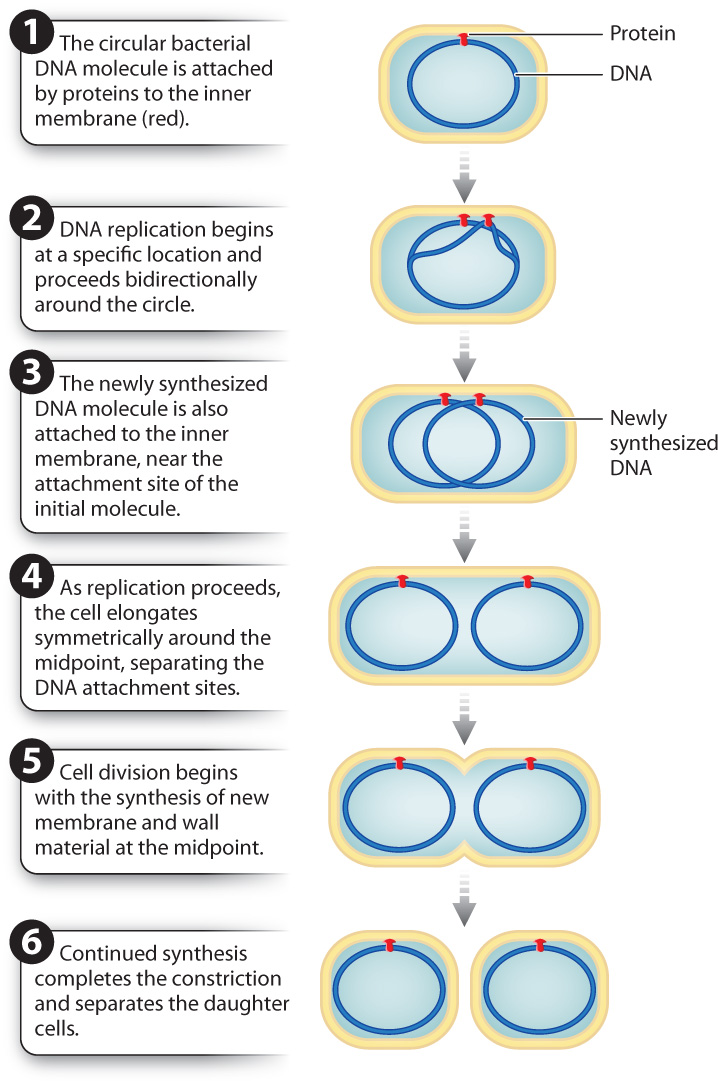
Like most cellular processes, binary fission requires the coordination of many components in both time and space. Recent research has identified several genes whose products play a key role in bacterial cell division. One of these genes, called FtsZ, has been especially well studied. The protein it encodes forms a ring at the site of constriction where the new cell wall forms between the two daughter cells. FtsZ is present in the genomes of diverse bacteria and archaeons, suggesting that it plays a fundamental role in prokaryotic cell division. Interestingly, it appears to be evolutionarily related to tubulin, which you will recall from Chapter 10 makes up the dynamic microtubules found in eukaryotic cells that are important in intracellular transport, cell movement, and cell division.
Question Quick Check 1
Z1frhISHaz+iwQIVeC52mUpw3c1jZv5vGeSER2i+RZgsTL7JFWRpsOPZFzcK/Bs/UjI2/hqTC06NKdMw2bmp7JnvyongGovy1x8PcaszgWgbc94Cq6MdXD5M5BoL5ALyuQJ2tx+Ru9IrcGJFBEtu6jTV67mrfYsjQd2ChErTTDo+fjfdvnGoMA==11-3
11.1.2 Eukaryotic cells reproduce by mitotic cell division.
The basic steps of binary fission that we just saw—replication of DNA, segregation of replicated DNA to daughter cells, and division of one cell into two—occur in all forms of cell division. However, cell division in eukaryotes is more complicated than cell division in prokaryotes. Instead of the single, relatively small, circular DNA molecule that is the genome of prokaryotic cells, the genome of eukaryotic cells is typically much larger and organized into one or more linear chromosomes, each of which needs to be replicated and separated into daughter cells. And whereas the DNA of prokaryotes is attached to the inside of the plasma membrane, allowing separation of replicated DNA into daughter cells by cell growth, the DNA of eukaryotes is located in the nucleus. As a result, eukaryotic cell division requires first the breakdown and then the re-formation of the nuclear envelope, as well as mechanisms other than cell growth to separate replicated DNA. As we saw in Chapter 10 and discuss in more detail in section 11.2, chromosomes of dividing eukaryotic cells attach to the mitotic spindle, which separates them into daughter cells.
Interestingly, some unicellular eukaryotes exhibit forms of cell division that have characteristics of binary fission and mitosis. For example, dinoflagellates, like all eukaryotes, have a nucleus and linear chromosomes. However, unlike most eukaryotes, the nuclear envelope does not break down but stays intact during cell division. Furthermore, the replicated DNA is attached to the nuclear envelope. The nucleus then grows and divides in a manner reminiscent of binary fission. These and observations of intermediate forms of cell division in other organisms strongly suggest that mitosis evolved from binary fission.
11.1.3 The cell cycle describes the life cycle of a eukaryotic cell.
Cell division in eukaryotic cells proceeds through a number of steps that make up the cell cycle (Fig. 11.2). The cell cycle consists of two distinct stages: M phase and interphase. During M phase, the parent cell divides into two daughter cells. M phase consists of two different events: (1) mitosis, the separation of the chromosomes into two nuclei, and (2) cytokinesis, the division of the cell itself into two separate cells. Usually, these two processes go hand in hand, with cytokinesis usually beginning even before mitosis is complete. In most mammalian cells, M phase lasts about an hour.
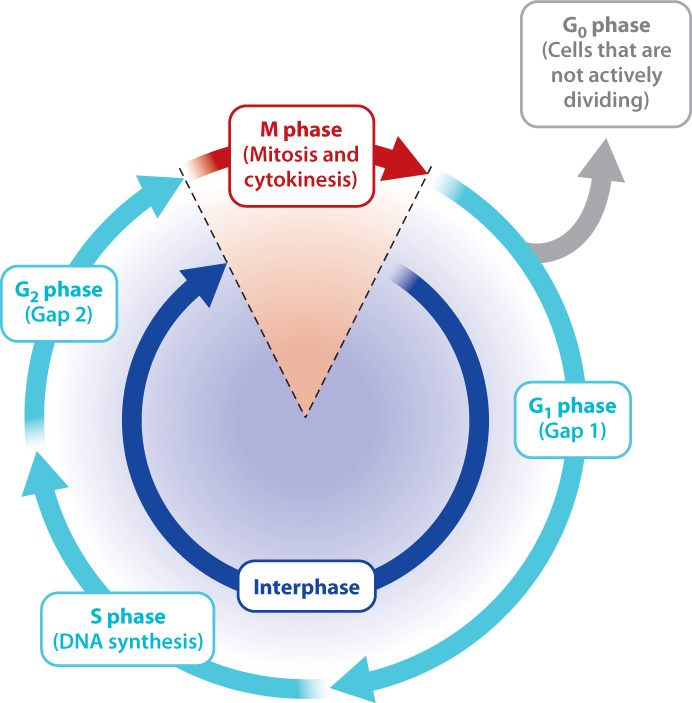
The second stage of the cell cycle, called interphase, is the time between two successive M phases (Fig. 11.2). This stage lasts about 10 to 14 hours in most actively multiplying mammalian cells. For many years, it was thought that the relatively long period of interphase is uneventful. Today we know that during this stage the cell makes many preparations for division. These preparations include replication of the DNA in the nucleus so that each daughter cell receives a copy of the genome, and an increase in cell size so that each daughter cell receives sufficient amounts of cytoplasmic and membrane components to survive on its own.
Interphase can be divided into three phases, as shown in Fig. 11.2. Among the many preparations that the cell must make during interphase, one particularly important task is the replication of the entire DNA content of the nucleus. Since replication involves the synthesis of DNA, this stage is called S phase (“S” standing for “synthesis”).
In most cells, S phase does not immediately precede or follow mitosis but is separated from it by two gap phases: G1 phase between the end of M phase and the start of S phase, and G2 phase between the end of S phase and the start of M phase. Many essential processes occur during both “gap” phases, despite the name. For example, both the size and protein content of the cell increase in preparation for division. In addition, the G1 phase is a time when specific regulatory proteins are made and activated. Once active, the regulatory proteins, many of which are kinases, then promote the activity of enzymes that synthesize DNA. Thus, G1 is a time of preparation for S-phase DNA synthesis, and G2 is a time of preparation for M-phase mitosis and cytokinesis.
The time it takes for cells to pass through the cell cycle depends on the type of cell and the organism’s stage of development. Actively dividing cells in some human tissues such as the intestine and skin require frequent replenishing. Cells in these tissues usually need about 12 hours to complete the cell cycle. Most other actively dividing cells in your body take about 24 hours to complete the cycle. A unicellular eukaryote like yeast can complete an entire cell cycle in just 90 minutes. Champions in the race through the cell cycle are the cells of the embryos of some frog species, in which early cell divisions divide the cytoplasm of the large egg cell into many smaller cells and so do not require any growth period in between cell divisions. Consequently, there are virtually no G1 and G2 phases and as little as 30 minutes pass between cell divisions.
11-4
Not all the cells in your body actively participate in the cell cycle, since not all tissues require rapid replenishing of cells. Instead, many cells pause in the cell cycle somewhere between M phase and S phase for periods ranging from days to more than a year. This period is called the G0 phase and is distinguished from G1 by the absence of preparations for DNA synthesis (Fig. 11.2). Although cells in G0 have exited the cell cycle, they are active in other ways, performing specialized functions. For example, liver cells remain in G0 for as much as a year, carrying out metabolism and detoxification, before reentering the cell cycle and dividing. Other cells such as those that form the lens of the eye and nerve cells enter G0 permanently and therefore are nondividing cells. Thus, many brain cells lost to disease or damage cannot be replaced.