11.2 MITOTIC CELL DIVISION
Mitotic cell division (mitosis followed by cytokinesis) is the normal mode of asexual reproduction in unicellular eukaryotes, and it is the means by which an organism’s cells, tissues, and organs develop and are maintained in multicellular eukaryotes. During mitosis and cytokinesis, the parental cell’s DNA is divided and passed on to two daughter cells. This process is continuous, but is divided into discrete steps marked by dramatic changes in the cytoskeleton and in the packaging and movement of the chromosomes.
11.2.1 The DNA of eukaryotic cells is organized as chromosomes.
One of the key challenges faced by a dividing eukaryotic cell is ensuring that both daughter cells receive an equal and complete set of chromosomes. The length of DNA contained in the nucleus of an average eukaryotic cell is on the order of 1 to 2 meters, well beyond the diameter of a cell. The DNA therefore needs to be condensed to fit into the nucleus, and then further condensed during cell division so that it does not become tangled as it segregates into daughter cells.
In eukaryotic cells, DNA is organized with histones and other proteins into chromatin, which can be looped and packaged to form the structures we know as chromosomes (Chapter 3). One of the earliest events in mitosis is the condensing of chromosomes from long, thin, threadlike structures typical of interphase to short, dense forms that are identifiable under the microscope during M phase.
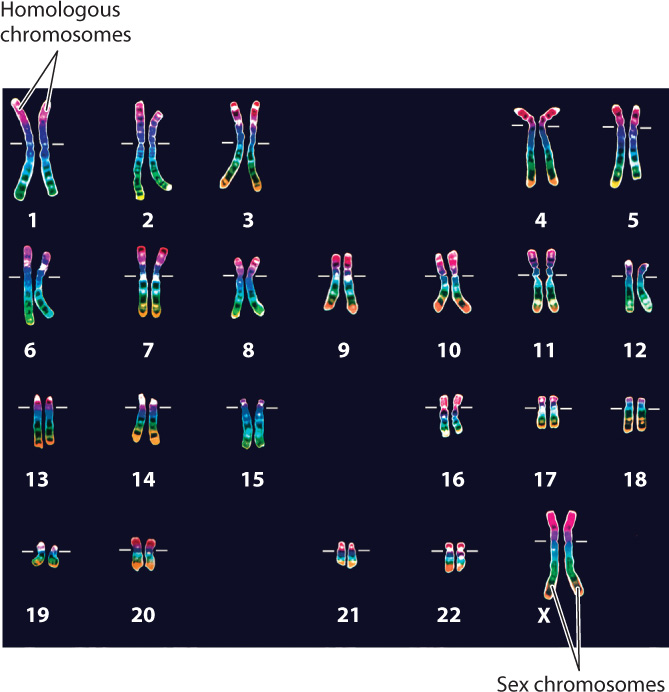
Every species is characterized by a specific number of chromosomes, and each chromosome contains a single molecule of DNA carrying a specific set of genes. When chromosomes condense and become visible during mitosis, they adopt characteristic shapes and sizes that allow each chromosome to be identified by its appearance in the microscope. The portrait formed by the number and shapes of chromosomes representative of a species is called its karyotype. Most of the cells in the human body, with the exception of the gametes, contain 46 chromosomes (Fig. 11.3). In contrast, cells from horses have 64 chromosomes, and cells from corn have 20.
11-5
In a normal human karyotype, the 46 chromosomes can be arranged into 23 pairs, 22 pairs of homologous chromosomes numbered 1 to 22 from the longest to the shortest chromosome and 1 pair of sex chromosomes (Fig. 11.3). Each pair of homologous chromosomes represents two of the same type of chromosome (both carrying the same set of genes), one of which was received from the mother and the other from the father. The sex chromosomes are the X and Y chromosomes. Individuals with two X chromosomes are female, and those with an X and a Y chromosome are male.
The number of complete sets of chromosomes in a cell is known as ploidy. A cell with one complete set of chromosomes is haploid, and a cell with two complete sets of chromosomes is diploid. Some organisms, such as plants, can have four or sometimes more complete sets of chromosomes. Such cells are polyploid.
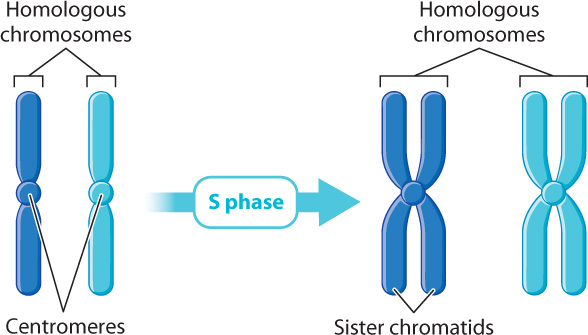
In order for cell division to proceed normally, every chromosome in the parent cell must be duplicated so that each daughter cell receives a full set of chromosomes. This duplication occurs during S phase. Even though the DNA in each chromosome duplicates, the two identical copies, called sister chromatids, do not separate. They stay side by side, physically held together at a constriction called the centromere. At the beginning of mitosis, the nucleus of a human cell contains 46 chromosomes, each of which is a pair of identical sister chromatids linked together at the centromere (Fig. 11.4). Thus, to count chromosomes, it is simply a matter of counting centromeres.
Question Quick Check 2
kQd7BruCWz1kxxqzgX9505BZDgWiykMtlalXkK/O2BY1qgIiIvtEQto1i7OgdT/vb4lj3X4rj+NbDz2znzTKpOdgx4K/As6AyX0F9+McC7uFT52vckWwaGLsHSwEWj2tPFabb8wuRiA=11.2.2 Prophase: Chromosomes condense and become visible.
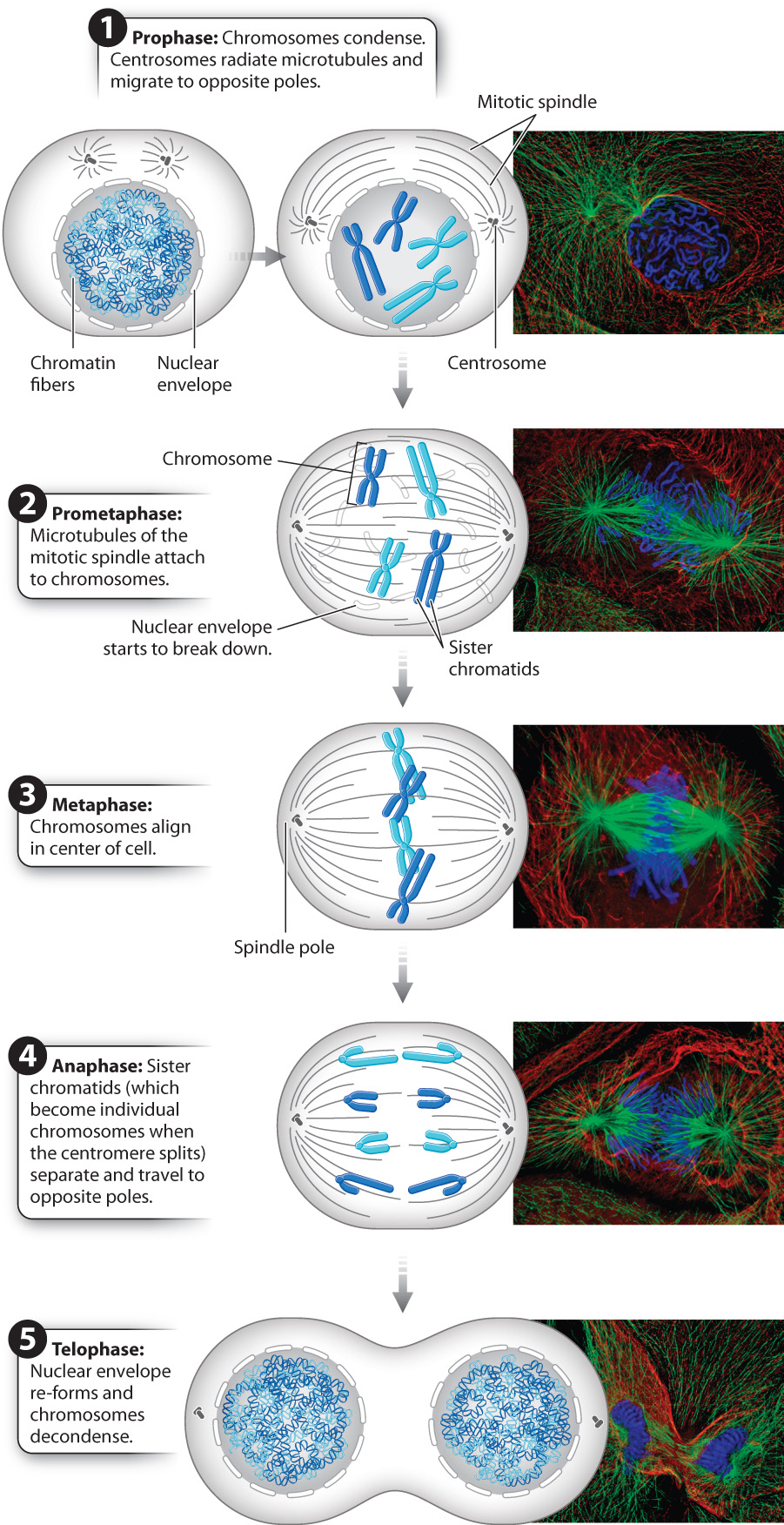
Mitosis takes place in five stages, each of which is easily identified by events that can be observed in the microscope (Fig. 11.5). When you look in a microscope at a cell in interphase, specific chromosomes cannot be distinguished because they are long and thin. As the cell moves from G2 phase to the start of mitosis, the chromosomes condense and become visible in the nucleus. The first stage of mitosis is known as prophase and is characterized by the appearance of visible chromosomes.
11-6
Outside the nucleus, in the cytosol, the cell begins to assemble the mitotic spindle, a structure made up predominantly of microtubules that pull the chromosomes into separate daughter cells. Recall from Chapter 10 that the centrosome is a compact structure that is the microtubule organizing center for animal cells. Plant cells also have microtubule-based mitotic spindles, but they lack centrosomes. As part of the preparation for mitosis during S phase in animal cells, the centrosome duplicates and each one begins to migrate around the nucleus, the two ultimately halting at opposite poles in the cell. The final locations of the centrosomes define the opposite ends of the cell that will eventually be separated into two daughter cells. As the centrosomes make their way to the poles of the cell, tubulin dimers assemble around them, forming microtubules that radiate from each centrosome. These radiating filaments form the mitotic spindle and later serve as the guide wires for chromosome movement.
11.2.3 Prometaphase: Chromosomes attach to the mitotic spindle.
In the next stage of mitosis, known as prometaphase, the nuclear envelope breaks down and the microtubules of the mitotic spindle attach to chromosomes (Fig. 11.5). The microtubules radiating from the centrosome poles grow and shrink as they explore the region of the cell where the nucleus once was. This process of growing and shrinking depends on the dynamic instability of microtubules, discussed in Chapter 10.
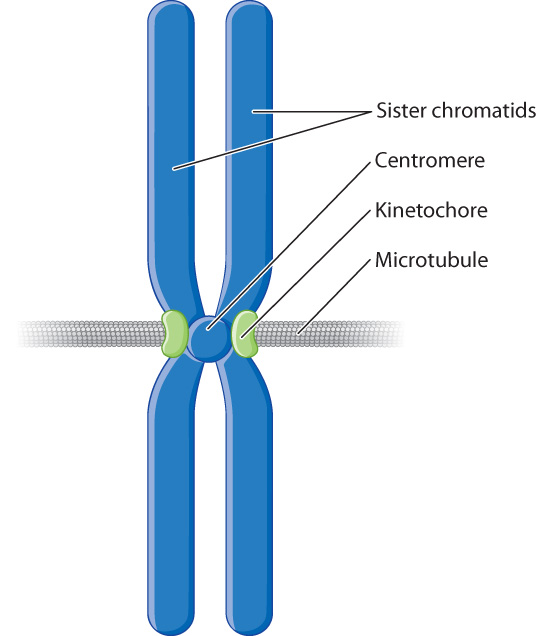
As the ends of the microtubules encounter chromosomes, they attach to the chromosomes at their centromeres. Associated with the centromere of each chromosome are two protein complexes called kinetochores, one located on each side of the constriction (Fig. 11.6). Each kinetochore is associated with one of the two sister chromatids and forms the site of attachment for a single microtubule. This arrangement ensures that each sister chromatid is attached to a microtubule radiating from one of the poles of the cell. The symmetrical tethering of each chromosome to the two poles of the cell is essential for proper chromosome segregation.
11.2.4 Metaphase: Chromosomes align as a result of dynamic changes in the mitotic spindle.
Once each chromosome is attached to both spindle poles, the microtubules of the mitotic spindle lengthen or shorten to move the chromosomes into position in the middle of the cell. There the chromosomes are lined up in a single plane that is roughly equidistant from both of the spindle poles. This stage of mitosis, when the chromosomes are aligned in the middle of the dividing cell, is called metaphase (see Fig. 11.5). It is one of the most visually distinctive stages under the microscope.
11.2.5 Anaphase: Sister chromatids fully separate.
In the next stage of mitosis, called anaphase, the sister chromatids separate (see Fig. 11.5). The centromere holding a pair of sister chromatids together divides, allowing the two sister chromatids to separate from each other. After separation, each chromatid is considered to be a full-fledged chromosome. The microtubules attached to the kinetochores gradually shorten, pulling the newly separated chromosomes to the opposite spindle poles of the cell.
This remarkable event is the reason for the equal segregation of chromosomes between the two daughter cells. In chromosome duplication during S phase in a human cell, each of the 46 chromosomes is duplicated to yield 46 pairs of identical sister chromatids. Thus, when the chromatids are separated at anaphase, an identical set of 46 chromosomes arrives at each spindle pole, the complete genetic material for one of the daughter cells.
11.2.6 Telophase: Nuclear envelopes re-form around newly segregated chromosomes.
Once a complete set of chromosomes arrives at a spindle pole, the chromosomes have entered the area that will form the cytosol of a new daughter cell. This event marks the beginning of telophase, during which cytosolic changes occur in preparation for the cell’s division into two new cells (see Fig. 11.5). The microtubules of the mitotic spindle break down and disappear, while nuclear envelopes begin to re-form around each set of chromosomes, creating two new nuclei. As the nuclei become increasingly distinct in the cell, the chromosomes contained within them decondense, becoming less visible in the microscope. This stage marks the end of mitosis.
11-7
11.2.7 The parent cell divides into two daughter cells by cytokinesis.
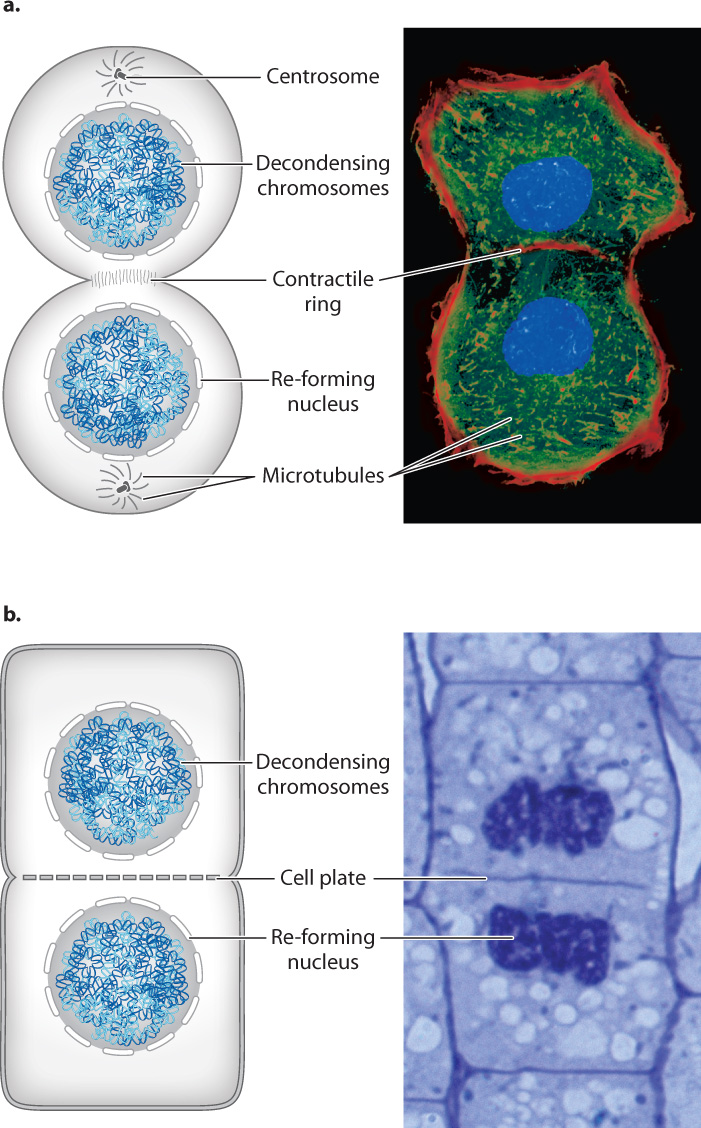
Usually, as mitosis is nearing its end, cytokinesis begins and the parent cell divides into two daughter cells (Fig. 11.7). In animal cells, this stage begins when a ring of actin filaments, called the contractile ring, forms against the inner face of the cell membrane at the equator of the cell perpendicular to the axis of what was the spindle (Fig. 11.7a). As if pulled by a drawstring, the ring contracts, pinching the cytoplasm of the cell and dividing it in two. This process is similar to what occurs in binary fission, though in the case of binary fission, the process is driven by FtsZ protein, a homolog of tubulin, not by actin as in eukaryotic cells. The constriction of the contractile ring is driven by motor proteins that slide bundles of actin filaments in opposite directions. Successful division results in two daughter cells, each with its own nucleus. The daughter cells are now free to enter G1 phase and start the process anew.
For the most part, mitosis is similar in animal and plant cells, but cytokinesis is different (Fig. 11.7b). Since plant cells have a cell wall, the division of the cell is achieved by constructing a new cell wall. During telophase, dividing plant cells form a structure called the phragmoplast in the middle of the cell. The phragmoplast consists of overlapping microtubules that guide vesicles containing cell wall components to the middle of the cell. During late anaphase and telophase, these vesicles fuse to form a new cell wall, called the cell plate, in the middle of the dividing cell. Once this developing cell wall is large enough, it fuses with the original cell wall at the perimeter of the cell. Cytokinesis is then complete and the plant cell has divided into two daughter cells.