13.4 ORGANIZATION OF GENOMES
The genomes of all organisms are large relative to the size of the cell. For example, if the circular genome of the intestinal bacterium Escherichia coli were fully extended, its length would be 200 times greater than the diameter of the cell itself. The fully extended length of DNA in human chromosome 1, our longest chromosome, would be 10,000 times greater than the diameter of the average human cell. There is consequently a need to package an enormous length of DNA into a form that will fit inside the cell while still allowing the DNA to replicate and carry out its coding functions. The mechanism of packaging differs substantially in bacteria, archaeons, and eukaryotes. We focus here on bacteria and eukaryotes, primarily because less is known about how DNA is packaged in archaeons.
13.4.1 Bacterial cells package their DNA as a nucleoid composed of many loops.
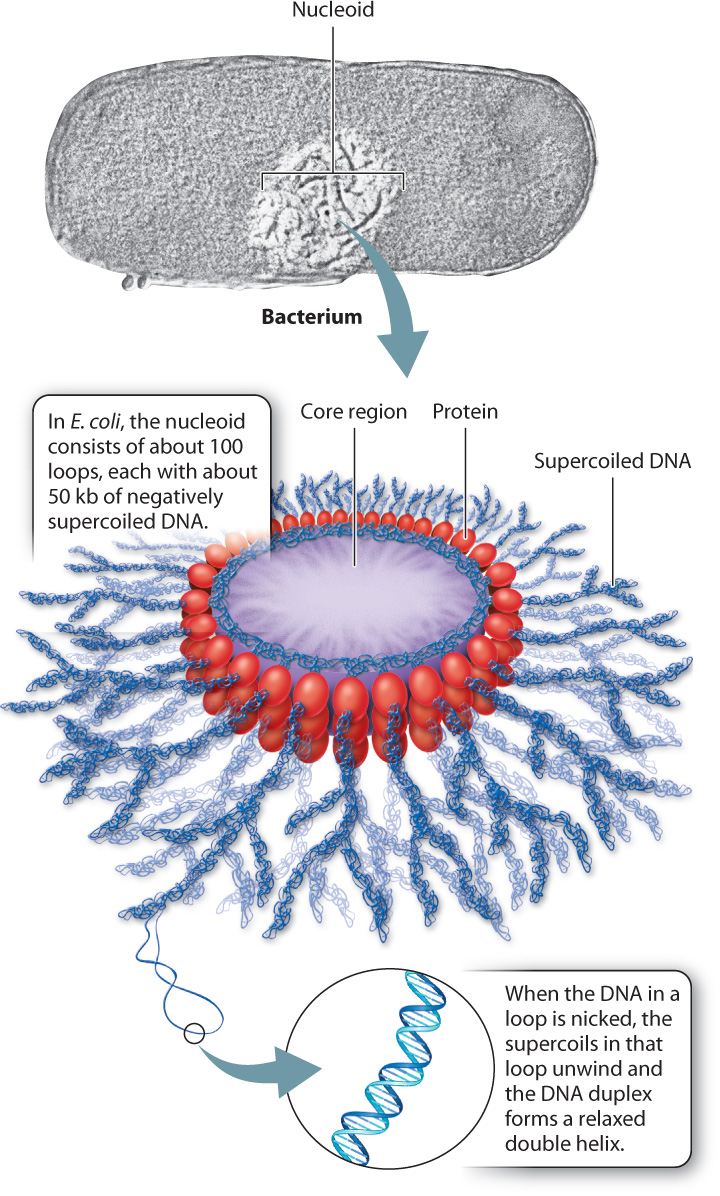
Bacterial genomes are circular, and the DNA double helix is underwound, which means that it makes fewer turns in going around the circle than would allow every base in one strand to pair with its partner base in the other strand. Underwinding is caused by an enzyme, topoisomerase II, that breaks the double helix, rotates the ends, and then seals the break. Underwinding creates strain on the DNA molecule, which is relieved by the formation of supercoils, in which the DNA molecule coils on itself. Supercoiling allows all the base pairs to form, even though the molecule is underwound. (You can make your own supercoil by stretching and twirling the ends of a rubber band, then relaxing the stress slightly to allow the twisted part to form coils around itself.) Supercoils that result from underwinding are called negative supercoils, and those that result from overwinding are positive supercoils. In most organisms, DNA is negatively supercoiled.
In bacteria, the supercoils of DNA form a structure with multiple loops called a nucleoid (Fig. 13.11). The supercoil loops are bound together by proteins. In E. coli, the nucleoid has about 100 loops, each containing about 50 kb of DNA. In addition to the protein binding that forms the loops, the negative supercoiling of the DNA serves to compress the molecule into a compact volume.
The supercoils in the loops of the nucleoid can be relaxed by introducing nicks, or interruptions, in the DNA backbones by means of enzymes or chemicals. The nicks allow the strands to rotate around each another to remove the supercoils, and the DNA molecule relaxes into the standard double helix. Any nick will relieve the supercoils in a loop, but only in the loop that is nicked. This means that each loop is supercoiled independently of the others (because of the proteins that hold the loops together).
13-12
13-13
13.4.2 Eukaryotic cells package their DNA as one molecule per chromosome.
In eukaryotic cells, DNA in the nucleus is packaged differently from that in bacteria (Fig. 13.12). Eukaryotic DNA is first wrapped around a group of histone proteins called a nucleosome (Chapter 3). Each nucleosome consists of two molecules, and each molecule consists of four different histone proteins. The histone proteins are rich in the amino acids lysine and arginine, whose positive charges neutralize the negative charges of the phosphates along the backbone of each DNA strand. This first level of packaging of the DNA is sometimes referred to as “beads on a string,” with the nucleosomes the beads, and the DNA the string.
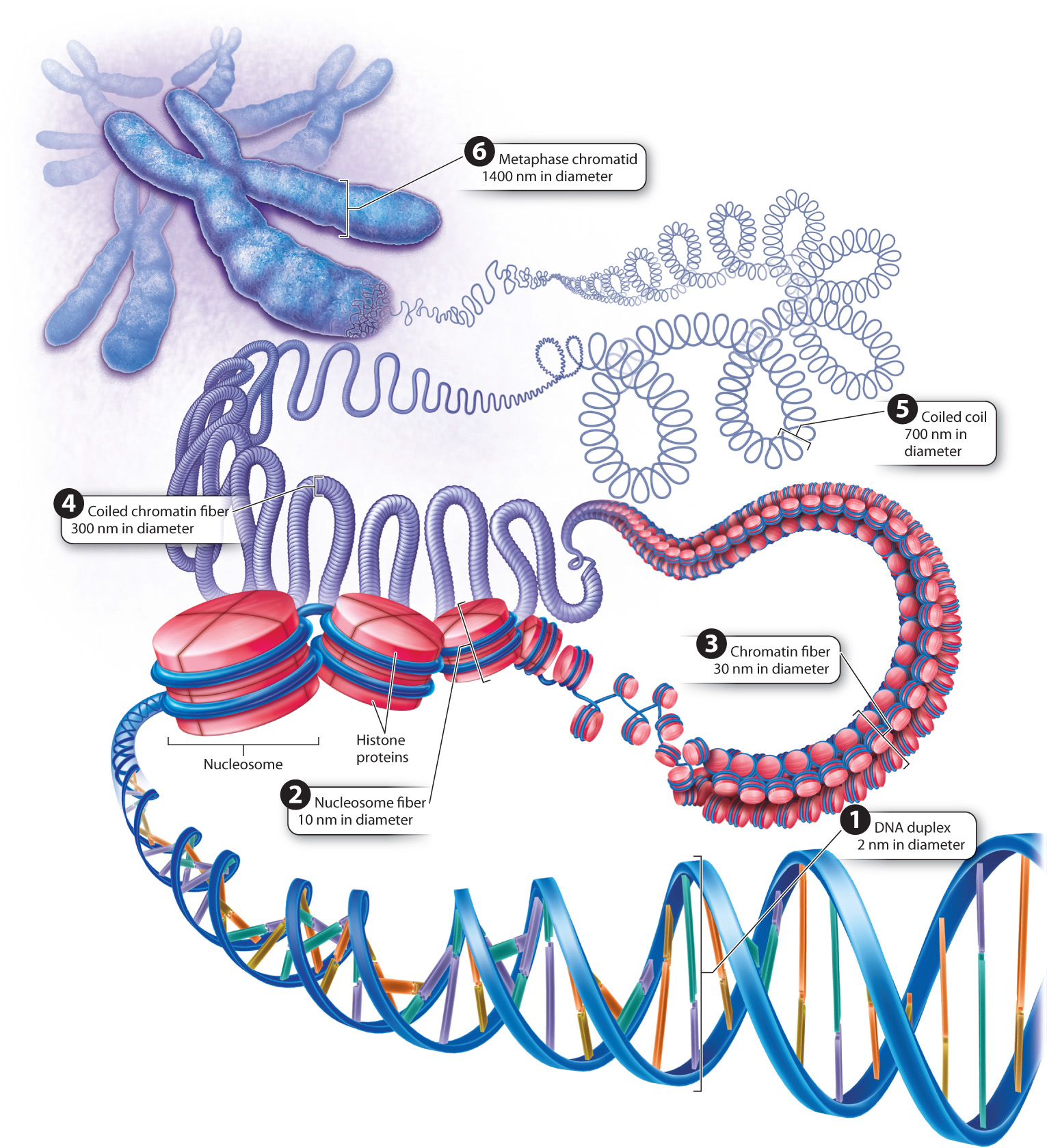
The nucleosomes and associated DNA are then coiled to form a structure called the 30-nm chromatin fiber (Fig. 13.12). When the chromosomes in the nucleus condense in preparation for cell division, each chromosome becomes progressively shorter and thicker as the 30-nm fiber coils in a manner that is still not fully understood. The progressive coiling constitutes chromosome condensation, an active, energy-consuming process requiring the participation of several types of proteins.
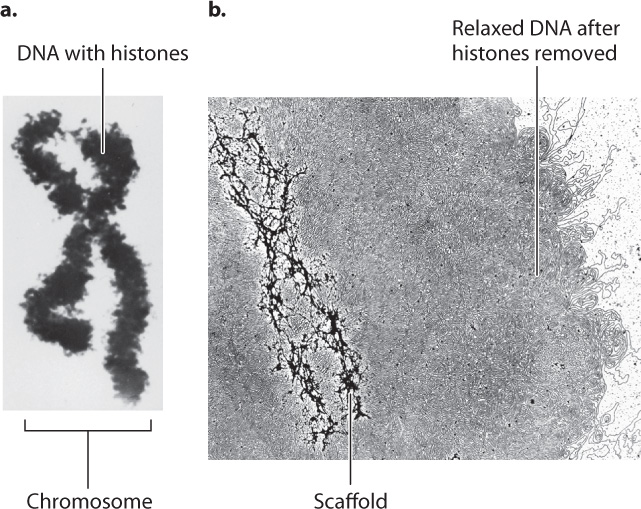
Greater detail of the structure of a fully condensed chromosome is revealed when the histones are chemically removed (Fig. 13.13). Without histones to coil around, the DNA spreads out in loops around a supporting protein structure called the chromosome scaffold. Each loop of relaxed DNA is 30 to 90 kb long and anchored to the scaffold at its base. Before removal of the histones, the loops are compact and supercoiled. Each human chromosome contains 2000–8000 such loops, depending on its size.
Despite intriguing similarities between the nucleoid model in Fig. 13.11 and the chromosome scaffold model in Fig. 13.13, the structures evolved independently and make use of different types of protein to bind the DNA and to form the folded structure of DNA and protein. Furthermore, the similarities are misleading, in part because the size of the eukaryotic chromosome is vastly greater than the size of the bacterial nucleoid. To appreciate the difference in scale, keep in mind that the volume of a fully condensed human chromosome is five times larger than the volume of a bacterial cell.
13.4.3 The human genome consists of 22 pairs of chromosomes and two sex chromosomes.
As emphasized in Chapter 11, the orderly process of meiosis is possible because chromosomes occur in pairs. The pairs usually match in size, general appearance, and position of the centromere, but there are exceptions, such as the X and Y sex chromosomes. The pairs of chromosomes that match in size and appearance are called homologous chromosomes. The members of each pair of homologous chromosomes have the same genes arranged in the same order along their length. If the DNA duplexes in each pair of homologs were denatured, each DNA strand could form a duplex with its complementary strand from the other homolog. There would be some differences in DNA sequence due to genetic variation, but not so many differences as to prevent DNA hybridization.
Chromosome painting illustrates the nearly identical nature of the DNA molecules in each pair of homologs. In this technique, individual chromosomes are isolated from cells in metaphase of mitosis. Metaphase of mitosis is the easiest stage in which to isolate chromosomes because of the availability of chemicals that prevent the spindle from forming. These chemicals block the cell cycle at metaphase, so cells progress to metaphase and then stop.
Once the chromosomes have been isolated, the DNA from each chromosome is fragmented, denatured, and labeled with a unique combination of fluorescent dyes. Under fluorescent light, the dyes give the DNA in each type of chromosome a different color. The fluorescently labeled DNA fragments are then mixed and hybridized to intact metaphase chromosomes from another cell. Each labeled fragment hybridizes to its complementary sequence in the metaphase chromosomes, “painting” each metaphase chromosome with dye-labeled DNA fragments.
13-14
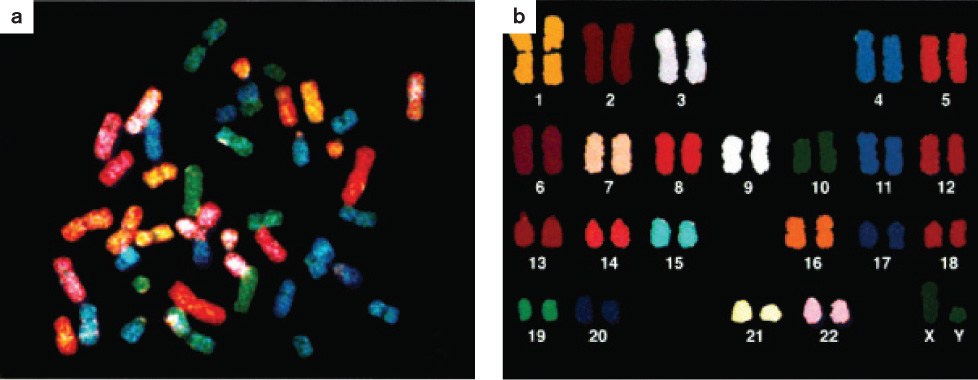
A chromosome paint of the chromosomes in a human male is shown in Fig. 13.14. Fig. 13.14a shows the chromosomes in the random orientation in which they were found in the metaphase cell, and Fig. 13.14b shows them arranged in a standard form called a karyotype (Chapter 11). To make a karyotype, the images of the homologous chromosomes are arranged in pairs from longest to shortest, with the sex chromosomes placed at the lower right. In this case, the sex chromosomes are XY (one X chromosome and one Y chromosome), indicating that the individual is male, whereas in a female the sex chromosomes would be XX (a pair of X chromosomes). Including the sex chromosomes, humans have 23 pairs of chromosomes.
An important observation from the chromosome paint shown in Fig. 13.14 is that the two members of each pair of homologous chromosomes show the same pattern of fluorescent color. This means that a particular labeled DNA fragment only hybridized with the two homologs of one chromosome. Hence, the DNA in each pair of homologous chromosomes is different from that in any other pair of homologous chromosomes.
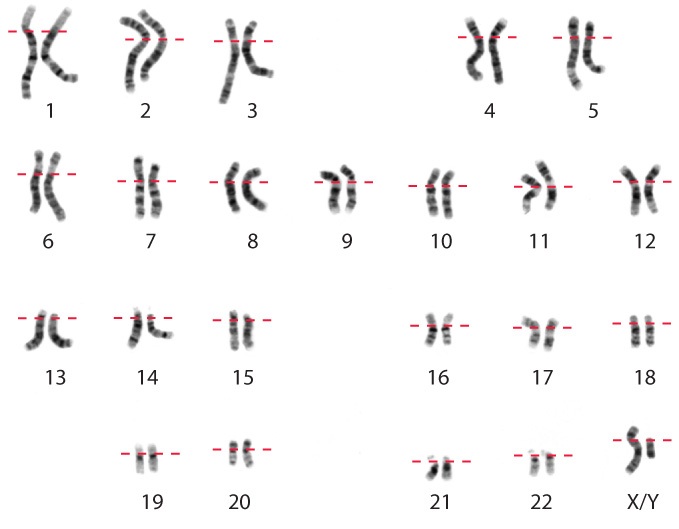
Higher resolution of human chromosomes can be obtained by the use of stains that bind preferentially to certain chromosomal regions and produce a visible pattern of bands, or crosswise striations, in the chromosomes. One such stain is the Giemsa stain; a Giemsa-stained karyotype is shown in Fig. 13.15. Note that each chromosome has a unique pattern of bands and that homologous chromosomes can readily be identified by their identical banding patterns. Procedures using the Giemsa stain yield about 300 bands that are used as landmarks for describing the location of genes along the chromosome.
Every species of eukaryote has its characteristic number of chromosomes. The number differs from one species to the next, with little relation between chromosome number and genome size. Despite variation in number, the rule that chromosomes come in pairs holds up pretty well, with the exception of the sex chromosomes. Polyploids, too, are an exception, but even in polyploids the number of copies of each homologous chromosome is usually an even number, so there are pairs of homologs after all.
The occurrence of chromosomes in pairs allows eukaryotes to reproduce sexually. When reproductive cells are formed during meiotic cell division, each cell receives one and only one copy of each of the pairs of homologous chromosomes (Chapter 11). When reproductive cells from two individuals fuse to form an offspring cell, the chromosome number characteristic of the species is reconstituted.
13.4.4 Organelle DNA forms nucleoids that differ from those in bacteria.
Most eukaryotic cells contain mitochondria, and many contain chloroplasts. Each type of organelle has its own DNA, meaning that eukaryotic cells have multiple genomes. Each has a nuclear genome consisting of the DNA in the chromosomes. Cells with mitochondria also have a mitochondrial genome, and those with chloroplasts also have a chloroplast genome.
Because the genome organization and mechanisms of protein synthesis in these organelles resemble those of bacteria, most biologists subscribe to the theory that the organelles originated as free-living bacterial cells that were engulfed by primitive eukaryotic cells billions of years ago (Chapter 27). In Chapters 7 and 8, we saw that the likely ancestor of mitochondria resembled a group of today’s non-photosynthetic bacteria (a group that includes E. coli), and the likely ancestor of chloroplasts resembled today’s photosynthetic cyanobacteria. In both cases, the DNA of the organelles became smaller during the course of evolution because most of the genes were transferred to the DNA in the nucleus. If the products of these transferred genes are needed in the organelles, they are synthesized in the cytoplasm and targeted for entry into the organelles by signal peptides (Chapter 5).
13-15
DNA in mitochondria and chloroplasts is often but not always circular, and there are usually multiple copies per organelle. Among animals, the size of a mitochondrial DNA molecule ranges from 14 kb to 18 kb. Mitochondrial DNA in plants is generally much larger, up to 100 kb. Plant chloroplast DNA is more uniform in size than mitochondrial DNA, with a range of 130 to 200 kb.
On the basis of size alone, some kind of packaging of organelle DNA is necessary. For example, the mitochondrial DNA in human cells is a circular molecule of 16 kb. Fully extended, it would have a circumference about as large as that of the mitochodrion itself. From their bacterial origin, you might expect organelle DNA to be packaged as a nucleoid rather than as a chromosome, and indeed it is. But the structures of the nucleoid in mitochondria and chloroplasts differ from each other, and also differ from those in free-living bacteria.