14.3 CHROMOSOMAL MUTATIONS
Whereas some mutations involve only one or a few nucleotides, some affect larger regions extending over thousands or millions of nucleotides, and a few have effects on chromosome structure that are large enough to be visible through a microscope. In this section, we briefly consider some of the major chromosome abnormalities. The most important are those in which the mutation affects the centromere, the site associated with attachment of the spindle fibers that move the chromosome during cell division (Chapter 11). An abnormal chromosome without a centromere or one with two centromeres is usually lost within a few cell divisions because it cannot be directed properly into the daughter cells during cell division.
14.3.1 Duplications and deletions result in gain or loss of DNA.
Among the most common chromosomal abnormalities are those in which a segment of the chromosome is either present in two copies or is missing altogether (Fig. 14.12). A chromosome in which a region is present twice instead of once is said to contain a duplication (Fig. 14.12a). Although large duplications that include hundreds or thousands of genes are usually harmful and quickly eliminated from the population, small duplications including only one or a few genes can be maintained over many generations. Usually, duplication of a region of the genome is less harmful than deletion of the same region.
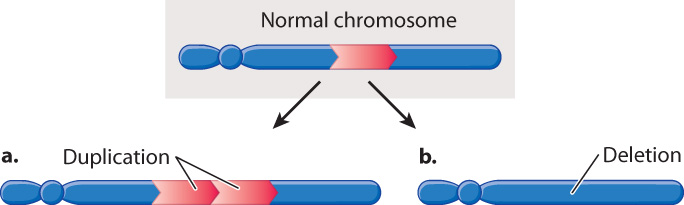
An example of a deletion, in which a region of the chromosome is missing, is shown in Fig. 14.12b. A deletion can result from an error in replication or from the joining of breaks in a chromosome that occur on either side of the deleted region. Even though a deletion may eliminate a gene that is essential for survival, the deletion can persist in the population because chromosomes usually occur in homologous pairs. If one member of a homologous pair has a deletion of an essential gene but the gene is present in the other member of the pair, that one copy of the gene is often sufficient for survival and reproduction. In these cases, the deletion can be transmitted from generation to generation and persist harmlessly, as long as the chromosome is present along with a normal chromosome.
But some deletions decrease the chance of survival or reproduction of an organism even when the homologous chromosome is normal. In general, the larger the deletion, the smaller the chance of survival. In the fruit fly Drosophila, individuals with deletions of more than 100–150 genes rarely survive even when the homologous chromosome is normal. The interpretation of the reduced survival is that organisms are sensitive to the dosage, or number of copies, of each gene. Normal embryonic development requires that genes be present in a particular dosage. Although small deviations from normal gene dosage can be tolerated, as indicated by the survival of individuals containing small duplications or deletions, the cumulative effects of large deviations from normal gene dosage are incompatible with life. It is usually not the total number of copies of each gene that matters, but rather the number of copies of each gene relative to other genes. This explains why some plant populations can include polyploids with multiple complete sets of chromosomes (Chapter 13), but in the same species large duplications and deletions are lethal.
14-12
14.3.2 Gene families arise from gene duplication and evolutionary divergence.
Small duplications play an important role in the origin of new genes in the course of evolution. In most cases, when a gene is duplicated, one of the copies is free to change without causing harm to the organism because the other copy continues to carry out the normal function of the gene. Occasionally, a mutation in the “extra” copy of the gene may result in a beneficial effect on survival or reproduction, and gradually a new gene is formed from the duplicate. These new genes usually have a function similar to that of the original gene.
This process of creating new genes from duplicates of old ones is known as duplication and divergence. Multiple rounds of duplication and divergence can give rise to a group of genes with related functions known as a gene family. The largest gene family in the human genome has about 400 genes and encodes for proteins that detect odors. These proteins are structurally very similar, but differ in the region that binds small odor molecules. It is the diversity of the odorant binding sites that allows us to identify so many different smells.
Most gene families are not as large and diverse as that for odor detection. Fig. 14.13 shows the evolutionary origin of the family of globin genes, which in humans are spread across about 50 kb of chromosome 11. The globin gene family consists of five different genes that are expressed at various times during development (embryo, fetus, or adult). The two γ- (gamma-) globin polypeptides are nearly identical in amino acid sequence but expressed at different levels in the fetus, whereas the adult δ- and β-polypeptides differ somewhat more and are expressed at very different levels. The sequences are all similar enough to imply that the genes arose through duplication and divergence.
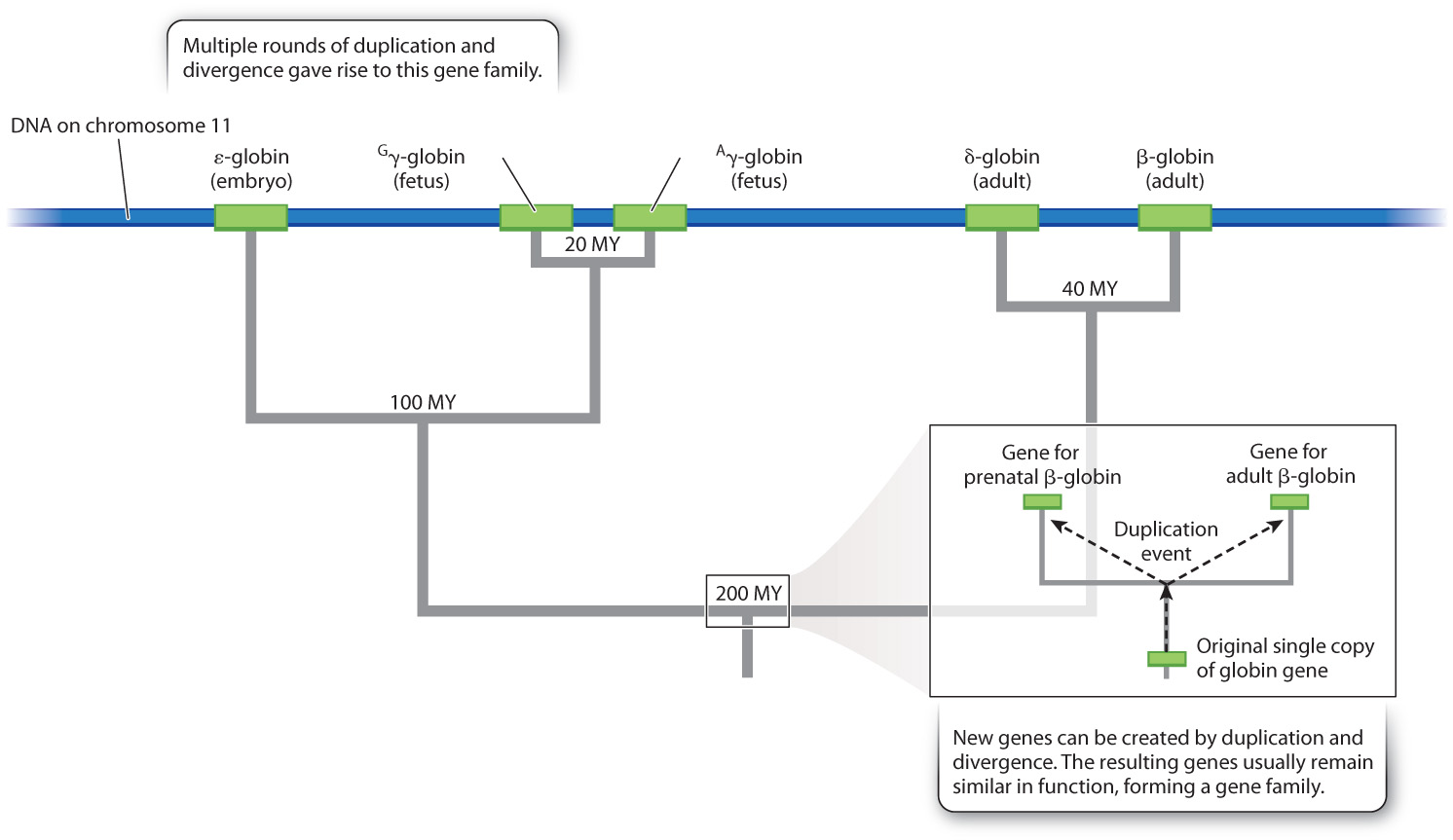
Globin sequences have changed through evolutionary time at a relatively constant rate, and so the fraction of differences between any two sequences is proportional to the time since they were created by duplication. The relative constancy of rates of evolutionary change in a DNA nucleotide sequence or a protein amino acid sequence is known as a molecular clock, and it allows molecular differences among genomes to be correlated with the fossil record and dated accordingly (Chapter 21). For example, the earliest duplication event in the tree in Fig. 14.13, which produced distinct genes for fetal and adult hemoglobin, took place at about the same time (200 million years ago) that the common ancestor of today’s placental mammals became a distinct species from the common ancestor of today’s marsupial mammals.
14.3.3 An inversion has a chromosomal region reversed in orientation.
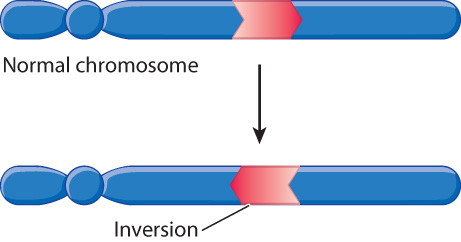
Chromosomes in which the normal order of a block of genes is reversed contain an inversion (Fig. 14.14). An inversion is typically produced when the region between two breaks in a chromosome is flipped in orientation before the breaks are repaired. Especially in large genomes, the breaks are likely to occur in noncoding DNA rather than within a gene. Whereas large inversions can cause problems in meiosis, small inversions are common in many populations and play an important role in chromosome evolution. The accumulation of inversions over evolutionary time explains in part why the order of genes along a chromosome can differ even among closely related species.
14-13
14.3.4 A reciprocal translocation joins segments from nonhomologous chromosomes.
A reciprocal translocation (Fig. 14.15) is said to have occurred when two different (nonhomologous) chromosomes undergo an exchange of parts. In the formation of a reciprocal translocation, both chromosomes are broken and the terminal segments are exchanged before the breaks are repaired. In large genomes, the breaks are likely to occur in noncoding DNA, so the breaks themselves do not usually disrupt gene function.
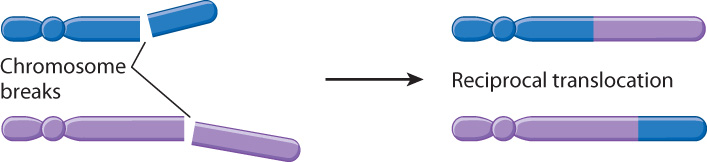
Since reciprocal translocations change only the arrangement of genes and not their number, most reciprocal translocations do not affect the survival of organisms. Proper gene dosage requires the presence of both parts of the reciprocal translocation, however, as well as one copy of each of the normal homologous chromosomes. Problems can arise in meiosis because both chromosomes involved in the reciprocal translocation may not move together into the same daughter cells, resulting in gametes with only one part of the reciprocal translocation. This inequality does upset gene dosage, because these gametes have extra copies of genes in one of the chromosomes and are missing copies of genes in the other. In the next chapter, we will see that such abnormalities are observed in a significant number of human embryos.