16.2 THE FOUNDATIONS OF MODERN TRANSMISSION GENETICS
Mendel’s scientific fame rests on the pea-breeding experiments that he carried out in the years 1856 to 1864 (Fig. 16.2), in which he demonstrated the basic principles of transmission genetics. His experiments were so simple and presented with such clarity that they still serve as examples of the scientific method at its best (Chapter 1).

16.2.1 Mendel’s experimental organism was the garden pea.
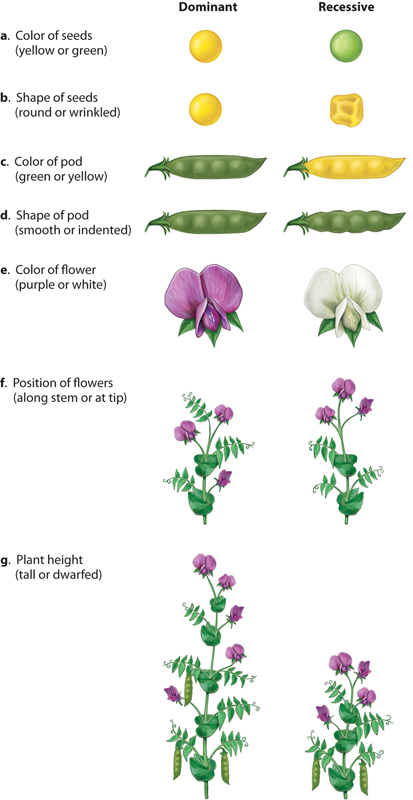
For his experimental material, Mendel used several strains of ordinary garden peas (Pisum sativum) that he obtained from a local seed supplier. His experimental approach was similar to that of a few botanists of the eighteenth and nineteenth centuries who studied the results of hybridization, or interbreeding between two different varieties or species of an organism. Where Mendel differed from his predecessors was in paying close attention to a small number of easily classified traits with contrasting characteristics. For example, where one strain of peas had yellow seeds, another had green seeds; and where one had round seeds, the other had wrinkled seeds (Fig. 16.3). Altogether Mendel studied seven physical features expressed in contrasting fashion among the strains: seed color, seed shape, pod color, pod shape, flower color, flower position, and plant height.
The expression of each trait in each strain was true breeding, which means that the physical appearance of the offspring in each successive generation is identical to the previous one. In other words, plants of the strain with yellow seeds produced only yellow seeds, and those of the strain with green seeds produced only green seeds. Likewise, plants of the strain with round seeds produced only round seeds, and those of the strain with wrinkled seeds produced only wrinkled seeds.
16-4
The objective of Mendel’s experiments was simple. By means of crosses between the true-breeding strains and crosses among their progeny, Mendel hoped to determine whether there are statistical patterns in the occurrence of the contrasting traits, such as yellow seeds or green seeds. If such patterns could be found, he would seek to devise a hypothesis to explain them and then use his hypothesis to predict the outcome of further crosses.
In designing his experiments, Mendel departed from other plant hybridizers of the time in three important ways:
- Mendel studied true breeding strains, unlike many other plant hybridizers who used complex and poorly defined material.
- Mendel focused on one trait, or a small number of traits, at a time, with characteristics that were easily contrasted among the true-breeding strains. Other plant hybridizers crossed strains differing in many traits and tried to follow all the traits at once. This resulted in amazingly complex inheritance, and no underlying patterns could be discerned.
- Mendel counted the progeny of his crosses, looking for statistical patterns in the offspring of his crosses. Others typically noted only whether offspring with a particular characteristic were present or absent, but did not keep track of and count all the progeny of a particular cross.
16.2.2 In crosses, one of the traits was dominant in the offspring.
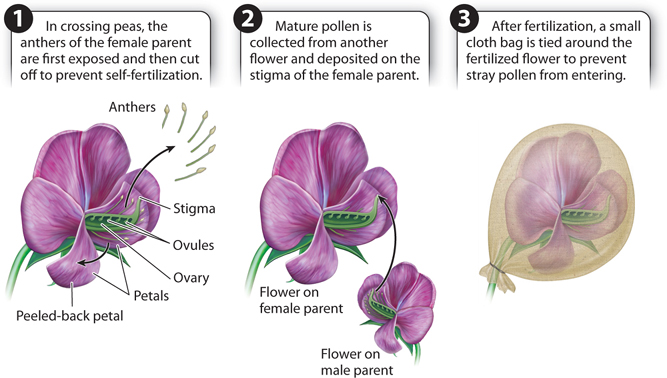
Mendel began his studies with crosses between true-breeding strains that differed in a single contrasting characteristic. Crossing peas is not as easy as it may sound. Pea flowers include both female and male reproductive organs enclosed together within petals (Fig. 16.4). Because of this arrangement, pea plants usually fertilize themselves (self-fertilize). Performing crosses between different plants is a tedious and painstaking process. First, the flower of the designated female parent must be opened at an early stage, and the immature anthers (plant male reproductive organs) clipped off and discarded so the plant cannot self-fertilize. Then mature pollen from the designated male parent must be collected and deposited on the stigma (female reproductive organ) of the female parent, from where it travels to the reproductive cells in the ovule at the base of the flower. And finally, a cloth bag must be tied around the female flower to prevent stray pollen from entering. No wonder Mendel complained that his eyes hurt!
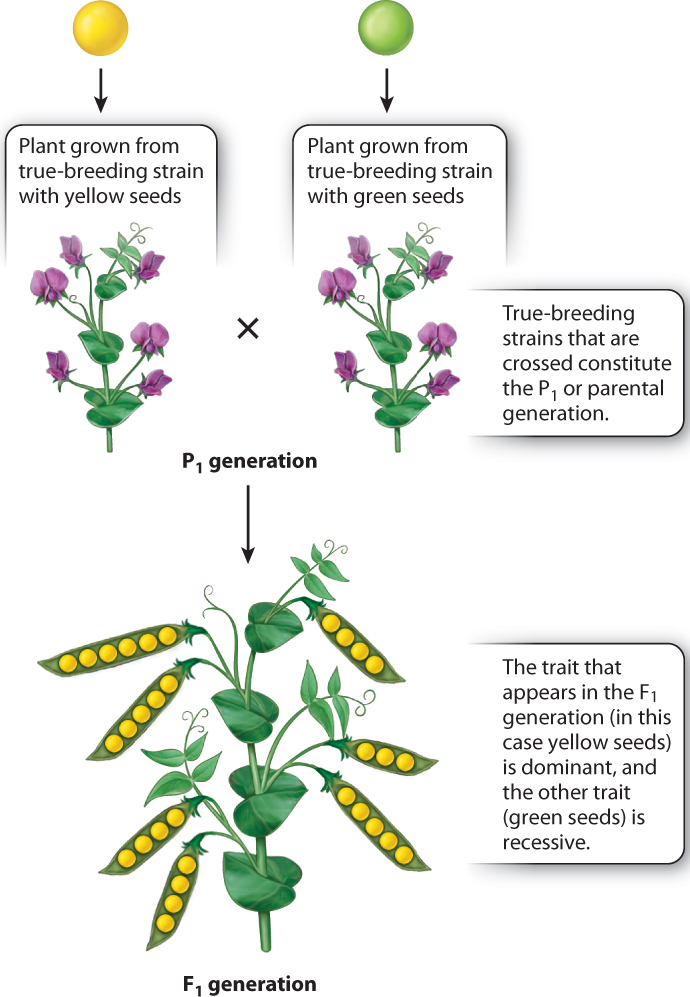
For each of the seven pairs of contrasting traits, true-breeding strains differing in the trait were crossed. Fig. 16.5 illustrates a typical result, in this case for yellow versus green seeds. In these kinds of crosses, the parental generation is referred to as the P1 generation, and the first offspring, or filial, generation, is referred to as the F1 generation. In the cross of P1 yellow × P1 green, Mendel observed that all the F1 progeny were yellow. This result was shown to be independent of the seed color, yellow or green, of the pollen donor because both of the crosses shown below yielded progeny plants with yellow seeds:

Crosses like these, in which the expressions of the trait in the female and male parents are interchanged, are known as reciprocal crosses. Mendel showed that reciprocal crosses yielded the same result for each of his seven pairs of contrasting traits.
Moreover, for each pair of contrasting traits, only one of the traits appeared in t he F1 generation. For these simple crosses, the trait that appears in the F1 generation is said to be dominant, and the contrasting trait that does not appear is said to be recessive. For each pair of traits illustrated in Fig. 16.3, the dominant trait is shown on the left, and the recessive on the right. Thus, yellow seed is dominant to green seed, round seed is dominant to wrinkled seed, and so forth. Mendel explained these findings by supposing that there is a hereditary factor for yellow seeds and a different hereditary factor for green seeds, likewise a hereditary factor for round seeds and a different one for wrinkled seeds, and so on. We now know that the hereditary factors that result in contrasting traits are different forms of a gene that affect the trait. The different forms of a gene are called alleles. The particular combination of alleles present in an individual constitutes its genotype, and the expression of the trait in the individual constitutes its phenotype.
16-5
In the cross between true-breeding plants with yellow seeds and true-breeding plants with green seeds, the genotype of each F1 seed includes an allele for yellow from the yellow parent and an allele for green from the green parent. The phenotype of each F1 seed is nevertheless yellow, because yellow is dominant to green.
The molecular basis of dominance is the fact that, in diploid organisms, only one copy of most genes is needed to carry out the normal function of the gene. For example, yellow pea seeds result from an enzyme that breaks down green chlorophyll, allowing yellow pigments to show through. Green seeds result from a mutation in this gene that inactivates the enzyme, and so the green chlorophyll is retained. In an F1 hybrid, such as that in Fig. 16.5, which receives a nonmutant gene from one parent and a mutant gene from the other, the seeds are yellow because one copy of the nonmutant gene produces enough of the enzyme to break down the chlorophyll to yield a yellow seed.