17.1 THE X AND Y SEX CHROMOSOMES
Among Mendel’s discoveries was that reciprocal crosses, in which the sexes of the parents associated with each genotype are reversed, yield the same types of progeny (Chapter 16). This principle is true of most genes, but an important exception applies to genes located in the X and Y chromosomes, which are associated with the sex of an individual. Mendel did not have to deal with this complication because peas and most other plants do not have sex chromosomes. In organisms that do have them, such as humans, specific rules govern the transmission of their genes.
17-2
17.1.1 In many animals, sex is genetically determined and associated with chromosomal differences.
Most chromosomes come in pairs that match in shape and size. The members of each pair are known as homologous chromosomes because they have the same arrangement of genes along their length (Chapter 11). One member of each pair of homologous chromosomes is inherited from the mother and the other from the father. In many animal species, however, the sex of an individual is determined by a special pair of unmatched chromosomes known as the sex chromosomes, which are usually designated as the X chromosome and the Y chromosome. Chromosomes other than the sex chromosomes are known as autosomes.
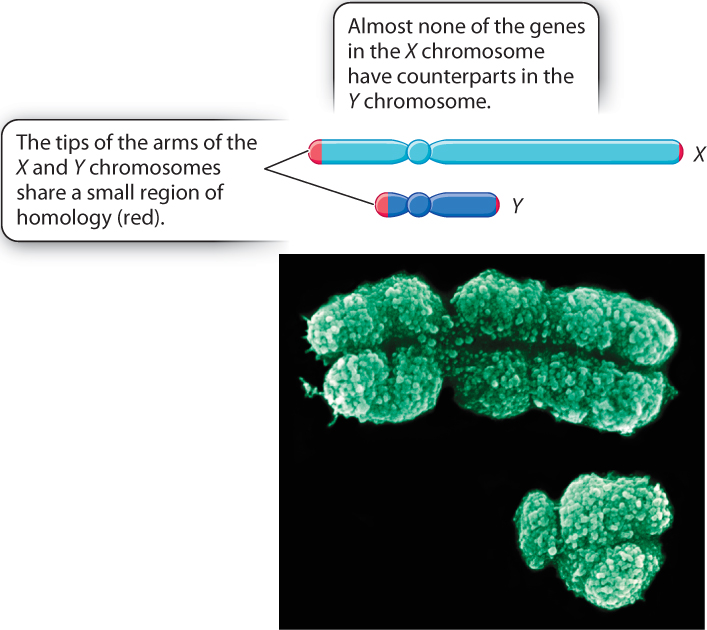
In humans, a normal female has two copies of the X chromosome (a sex-chromosome constitution denoted XX), and a normal male has one X chromosome and one Y chromosome (XY). The sizes of the human X and Y chromosomes are very different (Fig. 17.1). The X chromosome DNA molecule is more than 150 Mb long, while the Y chromosome is only about 50 Mb long. Except for a small region near each tip, the gene contents of the X and Y chromosomes are different from each other. The X chromosome, which includes more than 1000 genes, has a gene density similar to that of most autosomes. The vast majority of these genes have no counterpart in the Y chromosome. In contrast to the X chromosome, the Y chromosome contains only about 50 genes.
The regions of homology between the X and Y chromosomes consist of about 2.7 Mb of DNA near the tip of the short arm and about 0.3 Mb of DNA near the tip of the long arm. These regions of homology allow the chromosomes to pair during meiosis (Chapter 11). In most cells undergoing meiosis, a crossover (physical breakage, exchange of parts, and rejoining of the DNA molecules) occurs in the larger of these regions. The crossover allows the chromosomes to align correctly at metaphase I and to separate from each other at anaphase I.
The relative size and gene content of the X and Y chromosomes differ greatly among species. In some species of mosquitoes, the X and Y chromosomes are virtually identical in size and shape, and the regions of homology include almost the entire chromosome. This situation is unusual, however. In most species, the Y chromosome contains many fewer genes than the X chromosome. Some species, like grasshoppers, have no Y chromosome: Females in these species have two X chromosomes, and males have only one X chromosome. The total number of chromosomes in grasshoppers therefore differs between females and males.
17.1.2 Segregation of the sex chromosomes predicts a 1:1 ratio of females to males.
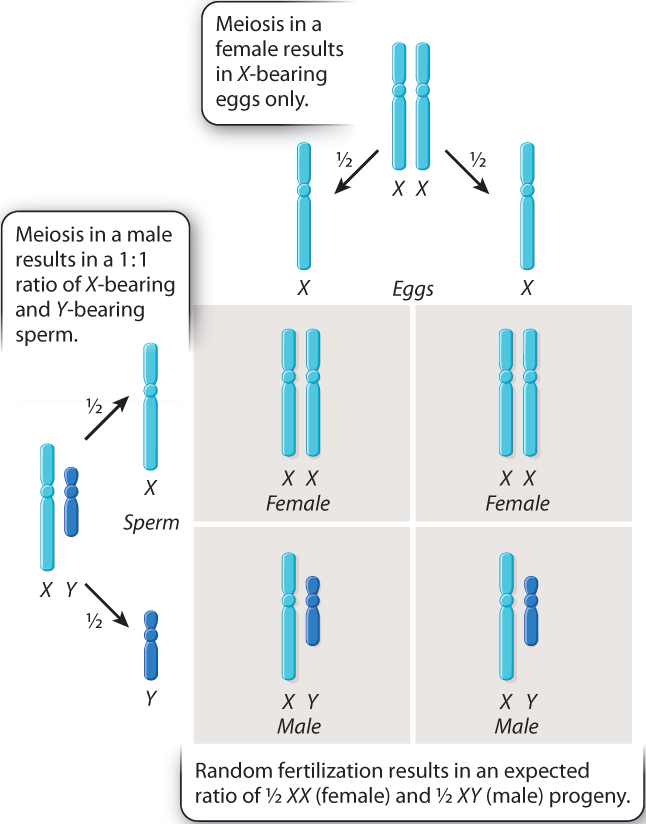
It is ironic that Mendel did not interpret sex as an inherited trait. If he had, he might have realized that sex itself provides one of the most convincing demonstrations of segregation. In human males, segregation of the X chromosome from the Y chromosome during anaphase I of meiosis results in half the sperm bearing an X chromosome and the other half bearing a Y chromosome (Fig. 17.2). Meiosis in human females results in eggs that each contain one X chromosome. With random fertilization, as shown in Fig. 17.2, half of the fertilized eggs are expected to be chromosomally XX (and therefore female) and half are expected to be chromosomally XY (and therefore male). This 1:1 ratio of female:male is characteristic of Mendelian segregation.
17-3
The expected ratio of 1:1 of females:males refers to the sex ratio at the time of conception. This is the primary sex ratio, and it is not easily observed because the earliest stages of fertilization and development are inaccessible to large-scale study. What can be observed is the sex ratio at birth, called the secondary sex ratio, which differs among populations but usually shows a slight excess of males. In the United States, for example, the secondary sex ratio is approximately 100 females:105 males. The explanation seems to be that, for reasons that are not entirely understood, females are slightly less likely to survive from conception to birth. On the other hand, males are slightly less likely to survive from birth to reproductive maturity, and so, at the age of reproductive maturity, the sex ratio is very nearly 1:1. Male mortality continues to be greater than that of females throughout life, so that by age 85 and over, the sex ratio is about 100 females:50 males.
The sex of each birth appears to be random relative to previous births—that is, there do not seem to be tendencies for some families to have boys or for other ones to have girls. Many people are surprised by this, for they know of one or more large families consisting mostly of boys or mostly of girls. But the occasional family with children of predominantly one sex would be expected simply by chance. When one occurs, its unusual sex distribution commands attention out of proportion to the actual numbers of such families. In short, families with unusual sex distributions are not more frequent than would be expected by chance.