21.4 NATURAL SELECTION
Natural selection results in allele frequencies changing from generation to generation according to the allele’s impact on the survival and reproduction of individuals. New mutations that are deleterious and eliminated by natural selection have no long-term evolutionary impact; ones that are beneficial, however, can result in adaptation to the environment over time.
21.4.1 Natural selection brings about adaptations.
The adaptations we see in the natural world—the exquisite fit of organisms to their environment—were typically taken by pre-Darwinian biologists as evidence of God’s existence. Each species, they argued, was so well adapted—the desert plant so physiologically adept at coping with minimal levels of rainfall and the fast-swimming fish so hydrodynamically streamlined—that it must have been deliberately designed by a divine Creator.

With the publication of On the Origin of Species in 1859, Darwin, pictured in Fig. 21.6, overturned the biological convention of his day on two fronts. First, he showed that species are not unchanging; they have evolved over time. Second, he suggested a mechanism, natural selection, that brings about adaptation. Natural selection was a brilliant solution to the central problem of biology: how organisms come to fit so well in their environments. From where does the woodpecker get its powerful chisel of a bill? And the hummingbird its long delicate bill for probing the nectar stores in flowers? Darwin showed how a simple mechanism, without foresight or intentionality, could result in the extraordinary range of adaptations that all of life is testimony to.
For 20 years after originally conceiving the essence of his theory, Darwin collected supporting evidence. In 1858, however, he was spurred to begin writing On the Origin of Species by a letter from a little-known naturalist collecting specimens in what is today Indonesia. By a remarkable coincidence, Alfred Russel Wallace, shown in Fig. 21.7, had also developed the theory of evolution by natural selection. Aware that Darwin was interested in the problem but having no idea that Darwin was working on the same theory, Wallace wrote to Darwin in 1858 to see what he thought of his idea.
Suddenly, Darwin was confronted with the prospect of losing his claim on the theory that he had been quietly nurturing for the previous 20 years. But all was not lost. Darwin’s colleagues arranged for the publication of a joint paper by Wallace and Darwin in 1858. This was done without consulting Wallace, who nonetheless never resented Darwin and afterward was careful to insist that the idea rightly belonged to the older man. It was Darwin’s publication of On the Origin of Species in 1859 that brought both evolution and its underlying mechanism of adaptation, natural selection, to public attention. Wallace is only fleetingly mentioned in Darwin’s great work, and Darwin is now the name, not Wallace, associated with the discovery.

Both Darwin and Wallace recognized their debt to the writings of a British clergyman, Thomas Malthus. In his Essay on the Principle of Population, first published in 1798, Malthus pointed out that natural populations have the potential to increase geometrically, meaning that there is potentially an accelerating increase in population size. Imagine that human couples can have just four children (two males and two females), meaning that the population doubles every generation. Starting with a single couple, by the twentieth generation, the population will have grown to over a million—1,048,576 to be precise. Note that this growth assumes a rather modest rate of increase (twofold). Consider how much faster population size would ramp up in an insect that lays hundreds of eggs in its lifetime.
21-9
However, this geometric expansion of populations does not occur. In fact, population sizes are typically reasonably stable from generation to generation. This is because the resources upon which populations are dependent—food, water, places to live—are limited. This imbalance suggests that in each generation many fail to survive or reproduce; there simply is not enough food and other resources to go around. This implies in turn that individuals within a population must compete for these resources.
Which individuals will win the competition? Darwin and Wallace suggested that those that are best adapted would most likely survive and leave more offspring. Genetic variation among individuals results in some being more likely to survive and reproduce. As a result, the next generation will be enriched for these same advantageous alleles. Darwin used the term “natural selection” for the filtering process that acts against deleterious alleles and in favor of advantageous ones.
Competitive advantage is a function of how well an organism is adapted to its environment. A desert plant that is more efficient at minimizing water loss than another plant is better adapted to the desert environment. An organism that is better adapted to its environment is more fit. By fit, we do not mean physically fit, or even optimally adapted. Fitness, in this context, is a measure of the extent to which the individual’s genotype is represented in the next generation. We say that the first plant’s fitness is higher than the second’s if it leaves more surviving offspring. Natural selection then acts over generations to increase the overall fitness of a population. A plant population newly arrived in a desert may be poorly adapted to its environment, but, over time, alleles that minimize water loss will increase under natural selection, resulting in a population that is better adapted to the desert.
Such changes in populations take time. Borrowing from the geologists of his day, Darwin recognized that time is a critical ingredient of his theory. Geologists had put forward a view of Earth’s history that argued that large geological changes—like the carving out of the Grand Canyon—can be explained by simple day-to-day processes—in this case, erosion—operating over vast timescales. Darwin applied this worldview to biology. He recognized that small changes, like subtle shifts in the frequencies of alleles, could add up to major changes given long enough time periods. What might seem to us to be a trivial change over the short term can, over the long term, result in substantial differences among populations.
21.4.2 The Modern Synthesis is a marriage between Mendelian genetics and Darwinian evolution.
Darwinian evolution involves the change over time of the genetic composition of populations and is thus a genetic theory. Although Mendel published his genetic studies of pea plants in 1866, not long after The Origin, Darwin never saw them, so a key component of the theory was missing. However, instead of clarifying the role of genetic processes underlying natural selection, the rediscovery of Mendel’s work in 1900 unexpectedly provoked a major controversy among evolutionary biologists. Some argued that Mendel’s discoveries did not apply to most genetic variation because the traits studied by Mendel were discrete, meaning that they had clear alternative states, such as peas that were either yellow or green. Most of the variation we see in natural populations, in contrast, is continuous, meaning that variation occurs across a spectrum. Human height, for example, does not come in discrete classes. People are not either 5 feet tall or 6 feet tall and no height in between. Instead, they may be any height within a certain range.
How could Mendel’s discrete traits account for the continuous variation seen in natural populations? This question was answered by the English theoretician Ronald Fisher, who realized that, instead of a single gene contributing to a trait like human height, there could be several genes that contribute to the trait. He argued that extending Mendel’s theory to include multiple genes per trait could account for patterns of continuous variation that we see all around us.
21-10
Fisher’s insight formed the basis of a synthesis between Darwin’s theory of natural selection and Mendelian genetics that was forged during the middle part of the twentieth century. The product of this Modern Synthesis was our current theory of evolution.
21.4.3 Natural selection increases the frequency of advantageous mutations and decreases the frequency of deleterious mutations.
Natural selection increases the frequency of beneficial alleles, resulting in adaptation. In some cases, it can promote the fixation of beneficial alleles, meaning the allele has a frequency of 1. To start with, a new beneficial allele will exist as a single copy in a single individual (that is, as a heterozygote), but, under the influence of natural selection, the beneficial allele can eventually replace all the other alleles in the population. Natural selection that increases the frequency of a favorable allele is called positive selection.
As we have seen, the majority of mutations to functional genes are deleterious. In extreme cases, they are lethal to the individuals carrying them and are thus instantly eliminated from the population. Sometimes, however, natural selection is inefficient in getting rid of a deleterious allele. Consider a recessive lethal mutation, b (that is, one that is lethal only as a homozygote, bb, and has no effect as a heterozygote, Bb). When it first arises, all the other alleles in the population are B, which means that the first b allele that appears in the population must be paired with a B allele, resulting in a Bb heterozygote. Because natural selection does not act against heterozygotes in this case, the b allele may increase in frequency by chance alone (we discuss how this happens below). Only when two b alleles come together to form a bb homozygote does natural selection act to rid the population of the allele. Natural selection that decreases the frequency of a harmful allele is called negative selection.
Many human genetic diseases show this pattern: The deleterious allele is rare and recessive. Because it is rare, homozygotes are formed only infrequently. Remember that the expected frequency of homozygotes under the Hardy–Weinberg equilibrium is the square of the frequency of the allele in the population. Therefore, if the allele frequency is 0.01, we expect 0.01 × 0.01, or 1 in every 10,000 individuals, to be homozygous for it. Thus the genetic disease occurs rarely, and the allele remains in the population because it is recessive and not expressed as a heterozygote.
21.4.4 What genetic differences have made some individuals more and some less susceptible to malaria?
Case 4 Malaria: Co-Evolution of Humans and a Parasite
In addition to allowing alleles to be either eliminated or fixed, natural selection can also maintain an allele at some intermediate frequency between 0% and 100%. This form of natural selection is called balancing selection, and it acts to maintain two or more alleles in a population. A simple case is members of a species that face different conditions depending upon where they live. One allele might be favored by natural selection in a dry area, but a different one favored in a wet area. Taking the species as a whole, these alleles are maintained by natural selection at intermediate frequencies.
Another example of balancing selection occurs when the heterozygote’s fitness is higher than that of either of the homozygotes, resulting in selection that ensures that both alleles remain in the population at intermediate frequencies. This form of balancing selection is called heterozygote advantage, and it is exemplified by human populations in Africa, where malaria has been a long-standing threat. Because the malaria parasite spends part of its life cycle in human red blood cells, mutations in the hemoglobin molecule that affect the structure of the red blood cells have a negative impact on the parasite and can reduce the severity of malarial attacks.
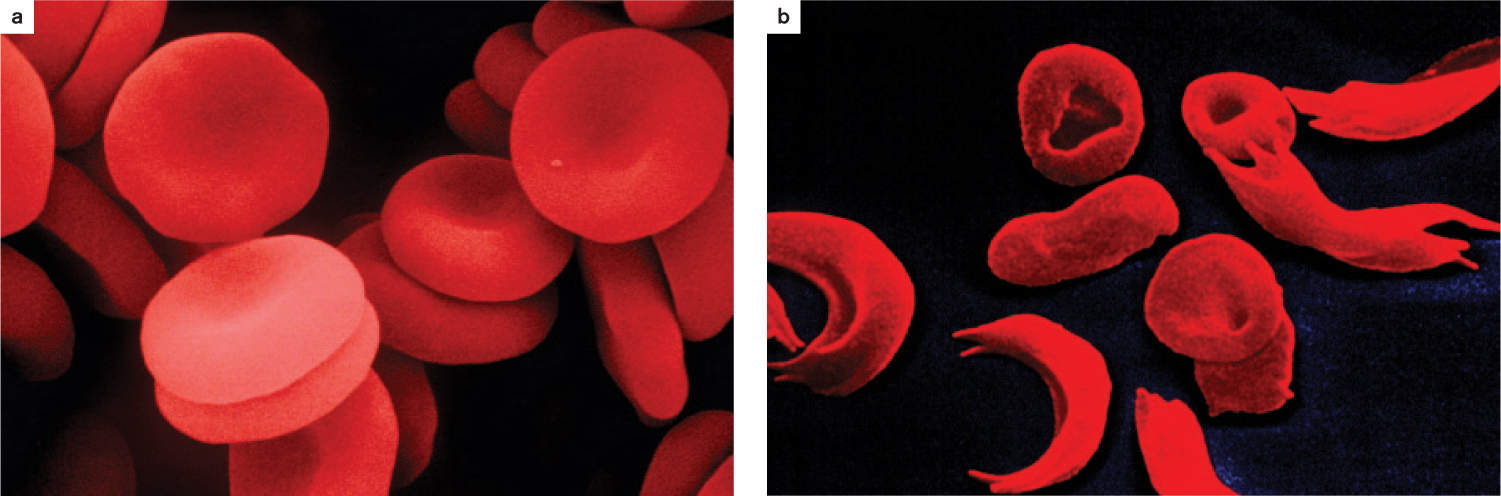
Two alleles of the gene for hemoglobin are A and S. The A allele codes for normal hemoglobin, resulting in fully functional, round red blood cells. The S allele causes the red blood cell to assume a sickle shape (Fig. 21.8). The S allele encodes a polypeptide that differs from the A allele’s product in just a single amino acid, which is enough to cause the molecules to aggregate end to end, so the red blood cell is distorted into a sickle (Chapter 15).
In regions of the world with malaria, heterozygous individuals (SA) have an advantage over homozygous individuals (SS and AA). SS homozygotes are protected against malaria, but they are burdened with severe sickling disease. Sickle-shaped red blood cells can block capillaries, and therefore people with two copies of S are prone to debilitating, painful, and sometimes fatal episodes resulting from capillary blockage. AA homozygotes lack sickling disease but are vulnerable to malaria. SA heterozygotes, however, do not have severe sickling disease and have some protection from malaria. As a result, natural selection maintains both the S and A alleles in the population at intermediate frequencies.
In areas where there is no malaria, this balance is shifted. Many African-Americans, descended from Africans upon whom natural selection operated in favor of the heterozygote, still carry the S allele, even though the allele is no longer useful to them in their malaria-free environment. If natural selection were to run its course among African-Americans, the S allele would gradually be eliminated. The problem is that this is a slow process, and many more people will continue to suffer from sickle cell anemia before it is complete.
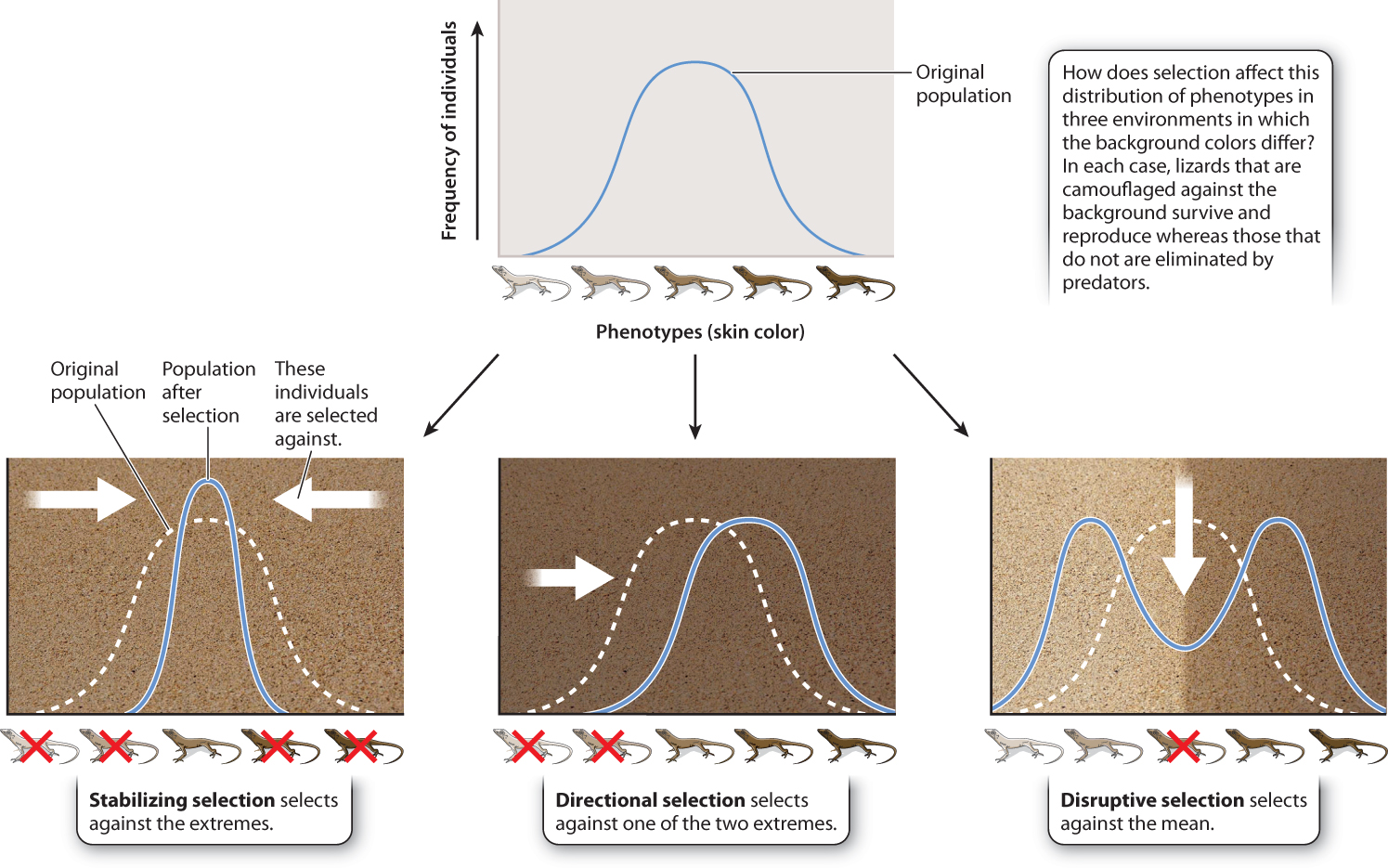
21.4.5 Natural selection can be stabilizing, directional, or disruptive.
Up to this point, we have followed the fate of individual mutations, which can increase, decrease, or be maintained at an intermediate frequency under the influence of natural selection. We can also look at the consequence of natural selection from a different perspective. Instead of following individual mutations, we can look at changes over time of a particular trait of an organism. For example we might track the evolution of height in a population, despite not knowing the specifics of the genetic basis of height differences. When we look at natural selection from this perspective, we see three types of patterns: stabilizing, directional, and disruptive, illustrated in Fig. 21.9.
21-11
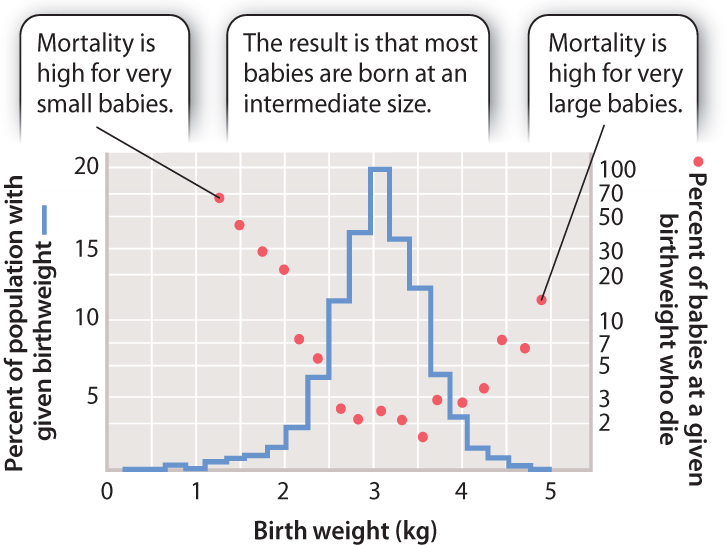
Stabilizing selection maintains the status quo and acts against extremes. A good example is provided by human birth weight, a trait affected by many factors, including many fetal genes (Fig. 21.10). If a baby is too small, then its chances of survival after birth are low. However, if it is too big, there may be complications that endanger both mother and baby during delivery. Thus, the optimum birth weight is between these two extremes. In this case, natural selection acts against the extremes. The vast majority of natural selection is of this kind as deleterious mutations that cause a departure from the optimal phenotype are selected against.
21-12
Whereas stabilizing selection keeps a trait the same over time, directional selection leads to a change in a trait over time. A timely and concerning example is provided by drug resistance in many viruses, bacteria, and the malaria parasite. In the case of malaria, widespread use of the drug chloroquine has led to changes in the population of the malaria parasite in favor of parasites that are resistant to this drug. From the point of view of drug resistance, the population has moved in one direction since the 1960s toward increased resistance.
Artificial selection, which has been practiced by humans since at least the dawn of agriculture, is a form of directional selection. Artificial selection is analogous to natural selection, but the competitive element is removed. Successful genotypes are selected by the breeder, not through competition. Because it can be carefully controlled by the breeder, artificial selection is astonishingly efficient at generating genetic change. Dogs are basically domesticated wolves that, over just a few millennia of careful breeding, have produced morphologies ranging from the tiny Pekingese to the vast Great Dane and behaviors ranging from the herding abilities of Border Collies to the retrieval skills of a Labrador.
Practiced over many generations, artificial selection can create a population in which the selected phenotype is far removed from that of the starting population. Fig. 21.11 shows the result of long-continued artificial selection for the oil content in kernels of corn.
FIG. 21.11: How far can artificial selection be taken?
BACKGROUND Begun in the 1890s and continuing to this day at the University of Illinois, this experiment is one of the longest-running biological experiments in history.
HYPOTHESIS Researchers hypothesized that there would be a limit to the extent to which the population could respond to continued directional selection, that there would be a point at which there was no longer a response to selection.
EXPERIMENT Corn was artificially selected for either high oil content or low oil content. Every generation, kernels showed a range of oil levels, but only the 12 kernels with the highest or the lowest oil content were used for the next generation.
RESULTS In the line selected for high oil content, the percentage of oil more than quadrupled, from about 5% to more than 20%. In the line selected for low oil content, the oil content fell so close to zero that it could no longer be measured accurately, and the selection was terminated. Both selected lines are completely outside the range of any phenotype observed at the beginning of the experiment.
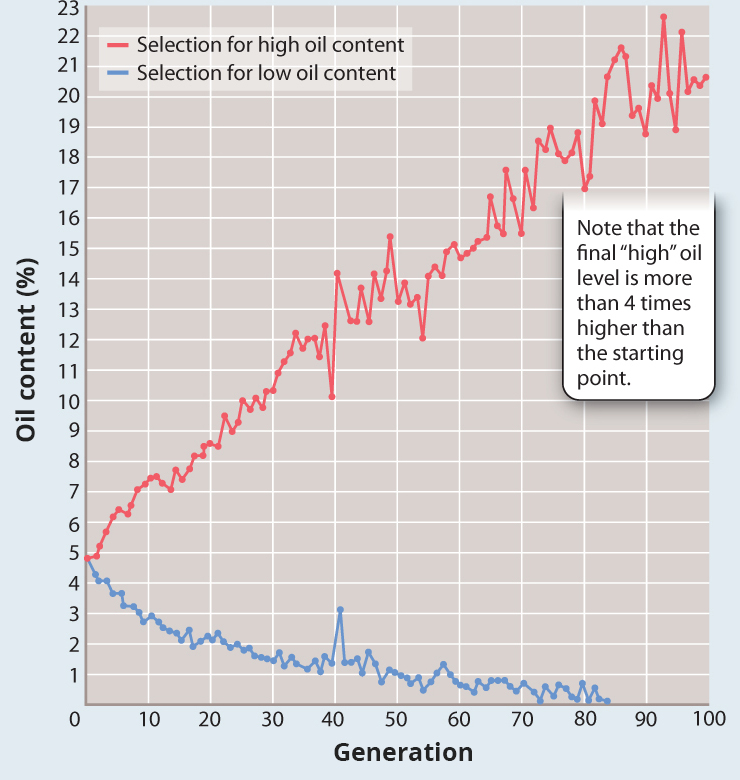
FOLLOW-UP WORK Genetic analysis of the selected lines indicates that the differences in oil content are due to the effects of at least 50 genes.
SOURCE Moose, S. P., J. W. Dudley, and T. R. Rocheford. 2004. “Maize Selection Passes the Century Mark: A Unique Resource for 21st Century Genomics.” Trends in Plant Science 9:358–364.
21-13
A third mode of selection, known as disruptive selection, operates in favor of extremes and against intermediate forms. For example, we may see disruptive selection operating on the size of seed-eating birds’ bills, with small (for delicate seeds) and large (for chunky seeds) bills favored over intermediate sizes that perform poorly with both delicate and chunky seeds. We explore this mechanism in more detail in the next chapter, as it can lead to the evolution of new species.
21.4.6 Sexual selection increases an individual’s reproductive success.
Initially, Darwin was puzzled by features of organisms that seemed to reduce an individual’s chances of survival. In a letter a few months after the publication of The Origin, he wrote, “The sight of a feather in a peacock’s tail, whenever I gaze at it, makes me sick!” The tail is metabolically expensive to produce; it is an advertisement to potential predators; and it is a massive encumbrance in any attempt to escape a predator. How could such a feature evolve under natural selection?
In his 1871 book, The Descent of Man, Darwin introduced a solution to this problem. Natural selection is indeed acting to reduce the showiness and size of the peacock’s tail, but another form of selection, sexual selection, is acting in the opposite direction. Sexual selection promotes traits that increase an individual’s access to reproductive opportunities. Peahens are thought to select their mates on the basis of the showiness and size of a male’s tail. In the absence of sexual selection, natural selection would act to minimize the size of the peacock’s tail. Presumably, the peacocks’ tails we see are a compromise, a trade-off between these conflicting demands of reproduction and survival (Chapter 45).