22.1 THE BIOLOGICAL SPECIES CONCEPT
The definition of species has been a long-standing problem in biology. Many biologists respond to the problem in the same way that Darwin himself did. In On the Origin of Species, Darwin wrote, “No one definition has as yet satisfied all naturalists; yet every naturalist knows vaguely what he means when he speaks of a species.” The difficulty of defining species has come to be called the species problem.
Here is the problem in a nutshell: The species, as an evolutionary unit, must by definition be fluid and capable of changing, giving rise through evolution to new species. The whole point of the Darwinian revolution is that species are not fixed. How, then, can we define something that changes over time, and, by the process of speciation, even leads to the origin of two species from one?
22.1.1 The species is the fundamental evolutionary unit.
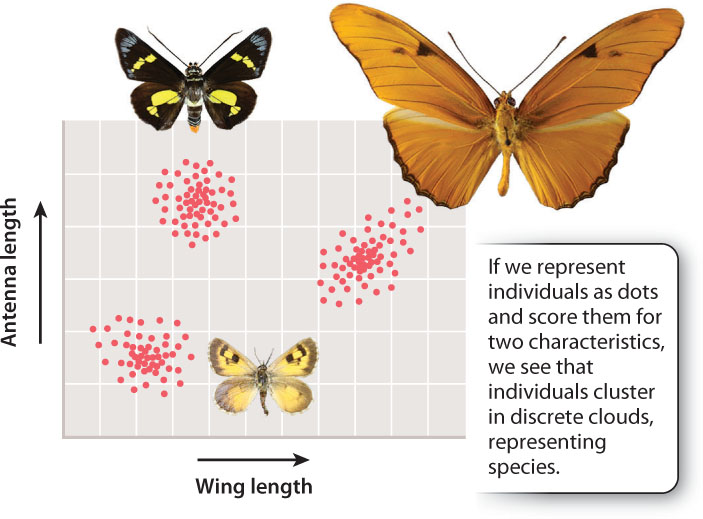
We can plainly see biodiversity, but are what we call “species” real biological entities? Or was the term coined by biologists to simplify their description of the natural world? If so, species would then be convenient categories that derive not from nature but from our desire to categorize what we see around us. To test whether or not species are real, we can examine the natural world, measure some characteristic of different living organisms we see, and then plot these measurements on a graph. Fig. 22.1 shows such a plot, graphing antenna length and wing length of three different types of butterfly. Note that the dots, representing individual organisms, fall into non-overlapping clusters. Each cluster is a species, and the fact that the clusters are distinct implies that species are biologically real. The distances we see between the dots within a cluster reflect variation from one individual to the next within a species (Chapter 21). Humans are highly variable, but overall we are more similar to one another than to our most humanlike relative, the chimpanzee. We form a messy cluster, but that cluster does not overlap with the chimpanzee cluster.
22-2
The species, then, is a fundamental biological unit. Taxonomists group similar species into genera (singular, genus), and similar genera into families, and so forth on the basis of shared ancestry. The lines between these higher taxonomic categories, however, are arbitrary; their composition depends on the judgment of taxonomists. If, in their view, two species are enough alike, they are placed in the same genus. If they are sufficiently different, they are placed in separate genera.
Whether or not two individuals are members of the same species, however, is not a matter of judgment, but rather a reflection of their ability (or inability) to exchange genetic material by producing fertile offspring. Furthermore, the species is the ultimate evolutionary unit. Think back to Chapter 21 and consider a new advantageous mutationthat appears initially in a single individual. That individual and its offspring inheriting the mutation will have a competitive advantage over other members of the population, and the mutation will increase in frequency until it reaches 100%. Migration among populations causes the mutation to spread further until all individuals within the species have it. The mutation spreads within the species but cannot spread beyond it. Therefore, the species is the key evolutionary unit. It is species that become extinct, and it is species that, through genetic divergence, give rise to new species.
22.1.2 Reproductive isolation is the key to the biological species concept (BSC).
Although biologists took until the middle part of the twentieth century to reach any kind of agreement on the definition of species, Alfred Russel Wallace, who discovered natural selection independently of Darwin, published a definition in 1865 that is almost identical to the one we use today:
Species are merely those strongly marked races or local forms which when in contact do not intermix, and when inhabiting distinct areas are generally believed to have had a separate origin, and to be incapable of producing a fertile hybrid offspring.
Wallace’s definition anticipates the key elements of the most widely used and generally accepted definition of a species, the biological species concept (BSC). The BSC was described by the great evolutionary biologist Ernst Mayr (1904–2005) as follows:
Species are groups of actually or potentially interbreeding populations that are reproductively isolated from other such groups.
Let us look at this definition closely. At the heart of it is the idea of reproductive compatibility. Members of the same species are capable of producing offspring together, whereas members of different species are incapable of producing offspring together. In other words, members of different species are reproductively isolated from one another. As Wallace and Mayr both realized, however, reproductive compatibility in fact entails more than just the ability to produce offspring. The offspring must be fertile and therefore capable of passing their genes on to their own offspring. For example, although a horse and a donkey, two different species, can mate to produce a mule, the mule is infertile. If, like a mule, a hybrid offspring is infertile, then it is a genetic dead end.
Note, too, the “actual or potential” part of Mayr’s definition. An Asian elephant living on the island of Sri Lanka and one living in nearby India are considered members of the same species, Elephas maximus, even though Sri Lankan and Indian Asian elephants never have a chance to mate with each other in nature because they are geographically separated (Fig. 22.2).

22.1.3 The BSC is more useful in theory than in practice.
Species, then, are real biological entities and the BSC is the most useful way to define them. However, the BSC has shortcomings, the most important of which is that it can be difficult to apply. Imagine you are on a field expedition in a rainforest and you find two insects that look reasonably alike. Are they members of the same species or not? To use the BSC, you would need to test whether or not they are capable of producing fertile offspring. However, in practice, you probably will not have the time and resources to perform such a test. And even if you do, it is possible that members of the same species will not reproduce when you place them together in a petri dish in your field camp. The conditions may be too unnatural or too threatening for the insects to behave in a normal way. And even if they do mate, you may not have time to determine whether or not the offspring are fertile. Wallace recognized this practical limitation of a reproduction-based definition of species, pointing out that in practice, “the test of hybridity cannot be applied in more than one case in ten thousand.”

22-3
Thus, we should consider the BSC as a valid framework for thinking about species, but one that is difficult to test. On a day-to-day basis, therefore, biologists often use a rule of thumb called the morphospecies concept. Stated simply, the morphospecies concept holds that members of the same species usually look alike, which is why natural history guidebooks are useful. For example, the shape, size, and coloration of the bald eagle make it easy to determine whether or not a bird observed in the wild is a member of the bald eagle species. Here we are returning to those biological clusters of Fig. 22.1 in which members of a species fall close to one another within a single, discrete group.
Although the morphospecies concept is a useful and generally applicable rule of thumb, it is not an infallible way to identify species. Members of a species may not always look alike, but instead show different phenotypes called polymorphisms (literally, “many forms”)—for example, color difference in some species of birds. In Chapter 21, we saw how minor genetic differences can result in two individuals within a species having very different appearances. Sometimes, males of a species may look different from females—think of the showy peacock, which differs dramatically from the relatively drab peahen. Young can be very different from old. Caterpillars that mature into butterflies are a striking example.
Whereas members of a species may look quite different from one another, the opposite can also be true: Members of different species can look quite similar. Some species of butterfly, for example, appear identical but have chromosomal differences that can be observed only with the aid of a microscope (Fig. 22.3). These chromosomal differences prevent the formation of interspecies hybrids, so those butterflies meet the BSC criterion—they are different species—but they do not meet the morphospecies criterion because they look so much alike.
22.1.4 The BSC does not apply to asexual or extinct organisms.
In addition to the difficulties of putting the BSC into practice, a second problem is that it overlooks some organisms. For example, because it is based on the sexual exchange of genetic information, the BSC cannot apply to species that reproduce asexually, as do many bacteria. Although some asexual species do occasionally exchange genetic information in a process known as conjugation (Chapter 26), true asexual organisms do not fit the BSC.
22-4
In addition, because it depends on reproduction, the BSC obviously cannot be applied to species that have become extinct and are known only through the fossil record, like different “species” of trilobites and dinosaurs.
22.1.5 Ring species and hybridization complicate the BSC.
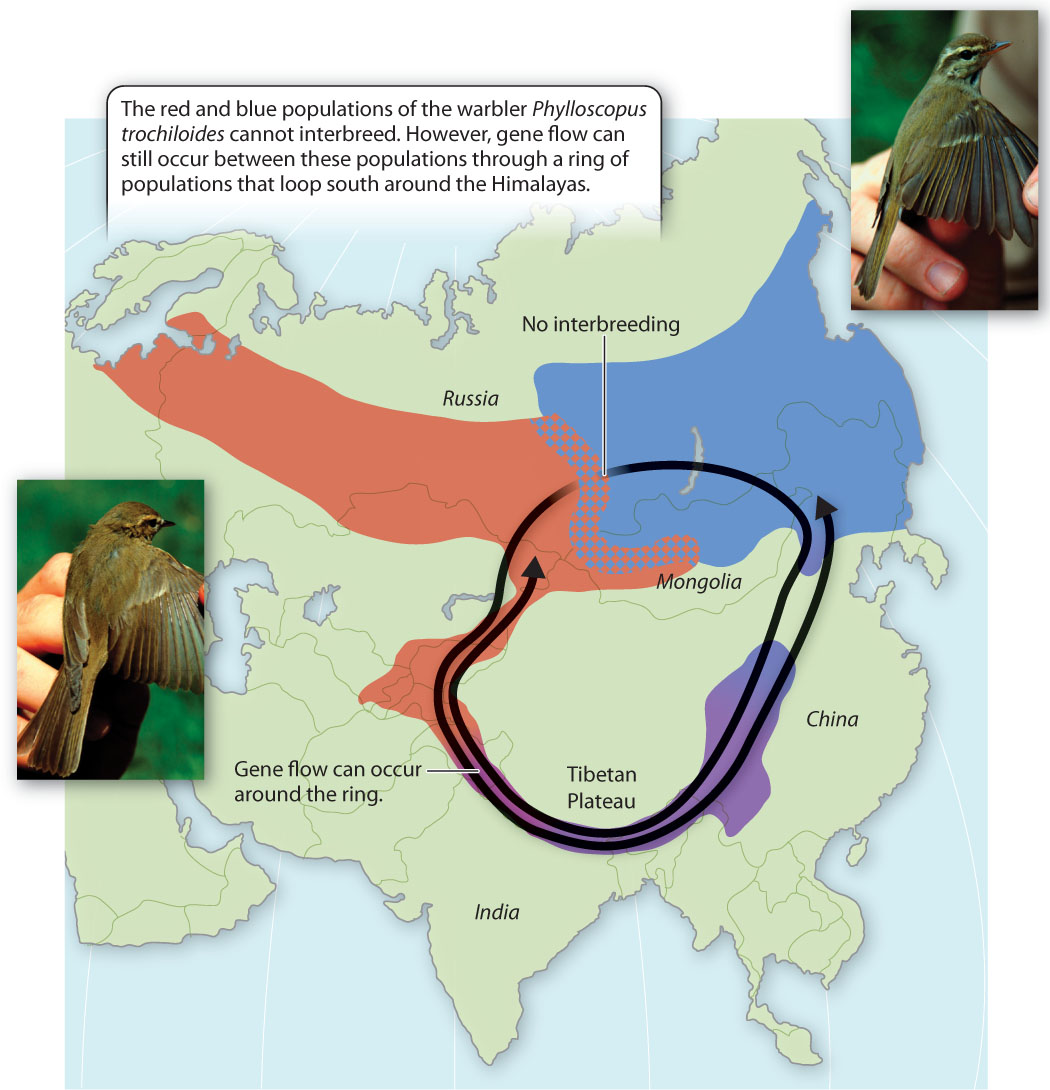
An unusual but interesting geographic pattern shown by ring species highlights another shortcoming of the BSC. Here we find that some populations within a species are reproductively isolated from each other, but others are not.
For example, populations of the greenish warbler, Phylloscopus trochiloides, are distributed in a large geographic loop around the Himalayas, as shown in Fig. 22.4. In Russia to the north, members of two neighboring populations do not interbreed. Therefore, according to the BSC, they are different species. However, the more western of the two Russian populations is capable of reproducing with the population to the south of it, and that population is capable of reproducing with the population to the east of it, and so on. Eventually, the loop of genetically exchanging populations comes all the way round to the more eastern of the two Russian populations. Thus, though members of the two Russian populations cannot exchange genes directly, they can do so indirectly, with the genetic material passing through many intermediate populations. This situation is more complicated than anything predicted by the BSC. The two Russian populations are reproductively isolated from each other but they are not genetically isolated from each other because of gene flow around the ring.
We also see a complicated situation in some groups of closely related species of plants. Despite apparently being good morphospecies that can be distinguished by appearance alone, many different species of willows (Salix), oaks (Quercus), and dandelions (Taraxacum) are still capable of exchanging genes with other species in their genera through hybridization, or interbreeding, between species. By the BSC, these different forms should be considered one large species because they are able to reproduce and produce fertile offspring. However, because they maintain their distinct appearances, natural selection must work against the hybrid offspring. This unusual phenomenon seems to occur mainly in plants.
22.1.6 Ecology and evolution can extend the BSC.
Because of these problems, much effort has been put into modifying and improving the BSC and many alternative definitions of species have been suggested. In general, these efforts highlight how difficult it is to make all species fit easily into one definition. The natural world truly defies neat categorization!
Several useful ideas, however, have come out of this literature. First is the notion that a species can sometimes be characterized by by its ecological niche, which, as we will discuss further in Chapter 47, is a complete description of the role the species plays in its environment—its habitat requirements, its nutritional and water needs, and the like. It turns out that it is impossible for two species to coexist in the same location if their niches are too similar because competition between them for resources will inevitably lead to the extinction of one of them. This observation has given rise to the ecological species concept (ESC), the idea that there is a one-to-one correspondence between a species and its niche. Thus, we can determine whether or not asexual bacterial lineages are distinct species on the basis of differences or similarities in their ecological requirements. If two lineages have very different nutritional needs, for example, we can infer on ecological grounds that they are separate species.
22-5
Second is the evolutionary species concept (EvSC), which is the idea that members of a species all share a common ancestry and a common fate. It is, after all, species rather than individuals that become extinct. The EvSC requires that all members of a species are descended from a single common ancestor. It does not specify, however, on what scale this idea should be applied. All mammals derive from a single common ancestor that lived about 200 million years ago, but there are thousands of what we recognize as species of mammals that have evolved since that long-ago common ancestor. But, under a strict application of the EvSC, would we consider all mammals to be a single species? Similarly, siblings and cousins are all descended from a common ancestor, a grandmother, but that is surely not sufficient grounds to classify more distant relatives as a distinct species. The EvSC can be useful when thinking about asexual species, but, given the arbitrariness of the decisions involved in assessing whether or not the descendants of a single ancestor warrant the term “species,” its utility is distinctly limited.
It is worth bearing these ecological and evolutionary considerations in mind when discussing and thinking about species. Although the BSC remains our most useful definition of species, the ESC and EvSC broaden and generalize the concept. For example, in the case of a normally asexual species that is difficult to study because conjugation is so infrequent, we can jointly apply the ESC and EvSC. We can use the ESC to loosely define the species in terms of its ecological characteristics (for example, its nutritional requirements), and we can refine that definition by using genetic analyses to determine whether the group is indeed a species by the EvSC’s standard (that is, that all its members derive from a single common ancestor).
Despite the shortcomings of the BSC and the usefulness of alternative ideas, we stress that the BSC is the most constructive way to think about species. In particular, by focusing on reproductive isolation—the inability to produce viable, fertile offspring—the BSC gives us a means of studying and understanding speciation, the process by which two populations, originally members of the same species, become distinct.