23.3 THE FOSSIL RECORD
Phylogenies based on living organisms provide hypotheses about evolutionary pattern. Branches toward the base of the tree occurred earlier than those near the top, and characters change and accumulate along the path from the root to the tips. Fossils provide direct documentation of ancient life, and so, in combination, fossils and phylogenies provide strong complementary insights into evolutionary history.
23.3.1 Fossils provide unique information.
Fossils can and do provide evidence for phylogenetic hypotheses, showing, for example, that organisms that branch early in phylogenies appear early in the geologic record. But the fossil record does more than this. First, fossils enable us to calibrate phylogenies in terms of time. It is one thing to infer that mammals diverged from the common ancestor of birds, crocodiles, lizards, and turtles before crocodiles and birds diverged from a common ancestor (see Fig. 23.7), but another matter to state that birds and crocodiles diverged about 220 million years ago, whereas the lineage represented today by mammals branched from other vertebrates about 100 million years earlier. As we saw in Chapter 21, estimates of divergence time can be made using molecular sequence data, but all such estimates must be calibrated using fossils.

The evolutionary relationship between birds and crocodiles highlights a second kind of information provided by fossils. Not only do fossils record past life, they also provide our only record of extinct species. The phylogeny in Fig. 23.7 contains a great deal of information, but it is silent about dinosaurs. Fossils demonstrate that dinosaurs once roamed Earth, and details of skeletal structure place birds among the dinosaurs in the vertebrate tree. Indeed, some remarkable fossils from China show that the dinosaurs most closely related to birds had feathers (Fig. 23.11).
23-12
A third, and also unique, contribution of fossils is that they place evolutionary events in the context of Earth’s dynamic environmental history. Again, dinosaurs illustrate the point. As discussed in Chapter 1, geologic evidence from several continents suggests that a large meteorite triggered drastic changes in the global environment 65 million years ago, leading to the extinction of most of the dinosaurs. In fact, at half a dozen moments in the past, large environmental disturbances sharply decreased Earth’s biological diversity. These events, called mass extinctions, have played a major role in shaping the course of evolution.
23.3.2 Fossils provide a selective record of past life.
Fossils are the remains of once-living organisms, preserved through time in sedimentary rocks. The feathered dinosaur shown in Fig. 23.11 is an example of one kind of fossil. If we wish to use fossils to complement phylogenies based on modern organisms, we must understand how fossils form and how the processes of formation govern what is and is not preserved.

For all its merits, the fossil record should not be thought of as a complete dictionary of everything that ever walked, crawled, or swam across our planet’s surface. Fossilization requires burial, as when a clam dies on the seafloor and is quickly covered by sand, or a leaf falls to the forest floor and ensuing floods cover it in mud. Through time, accumulating sediments harden into sedimentary rocks such as those exposed so dramatically in the walls of the Grand Canyon (Fig. 23.12). Without burial, the remains of organisms are eventually recycled by biological and physical processes, and no fossil forms. In general, the fossil record of marine life is more complete than that for land-dwelling creatures because marine habitats are more likely than those on land to be places where sediments accumulate and become rock. Thus, trees and elks living high in the Rocky Mountains have a low probability of fossilization, whereas clams and corals on the shallow seafloor are commonly buried and become fossils.
Biological factors contribute to the incompleteness of the fossil record. Most fossils preserve the hard parts of organisms, those features that resist decay after death. For animals, this usually means mineralized skeletons. Clams and snails that secrete shells of calcium carbonate have excellent fossil records. More than 80% of the clam species found today along California’s coast also occur as fossils in sediments deposited during the past million years. In contrast, nematodes, tiny worms that may be the most abundant animals on Earth, have no mineralized skeletons and almost no fossil record. The wood and pollen of plants, which are made in part of decay-resistant organic compounds, enter the fossil record far more commonly than do flowers. And, among unicellular organisms, the skeleton-forming diatoms, radiolarians, and foraminiferans have exceptionally good fossil records, whereas most amoebas are unrepresented.
23-13

Together, then, the properties of organisms (do they make skeletons or other features that resist decay after death?) and environment (did the organisms live in a place where burial was likely?) determine the probability that an ancient species will be represented in the fossil record. Fortunately, for species that make mineralized skeletons and live on the shallow seafloor, the fossil record is very good, preserving a detailed history of evolution through time.
Organisms that lack hard parts can leave a fossil record in two other, distinctive ways. Many animals leave tracks and trails as they move about or burrow into sediments. These trace fossils, from dinosaur tracks to the feeding trails of snails and trilobites, preserve a record of both anatomy and behavior (Fig. 23.13).
Organisms can also contribute molecular fossils to the rocks. Most biomolecules decay quickly after death. Proteins and DNA, for example, generally break down before they can be preserved, although, remarkably, a sizable fraction of the Neanderthal genome has been pieced together from DNA in 40,000-year-old bones. Other molecules, especially lipids like cholesterol, are more resistant to decomposition. Sterols, bacterial lipids, and some pigment molecules can accumulate in sedimentary rocks, documenting organisms that rarely form conventional fossils, especially bacteria and single-celled eukaryotes.
Rarely, unusual conditions preserve fossils of unexpected quality, including animals without shells, delicate flowers or mushrooms, fragile seaweeds or bacteria, even the embryos of plants and animals. For example, 505 million years ago, during the Cambrian Period, a sedimentary rock formation called the Burgess Shale accumulated on a relatively deep seafloor covering what is now British Columbia. Waters just above the basin floor contained little or no oxygen, so that when mud swept into the basin, entombed animals were sealed off from scavengers, disruptive burrowing activity, and even bacterial decay. For this reason, Burgess rocks preserve a remarkable sampling of marine life during the initial diversification of animals (Fig. 23.14).
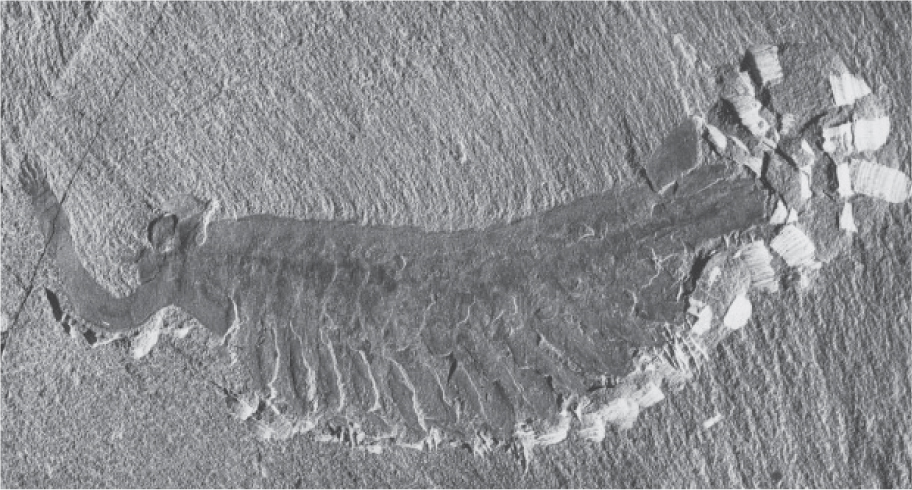
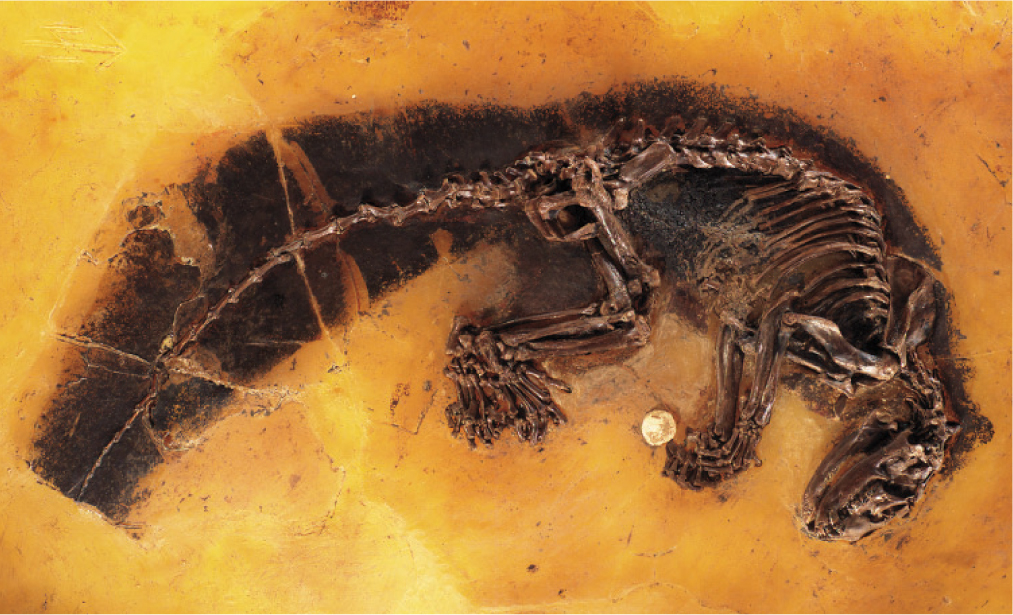
The Messel Shale formed more recently—about 50 million years ago—in a lake in what is now Germany. Release of toxic gases from deep within the Messel Lake suffocated local animals, and their carcasses settled into oxygen-poor muds on the lakefloor. Fish, birds, mammals, and reptiles are preserved as complete and articulated skeletons, and mammals retain impressions of fur and color patterning (Fig. 23.15). Plants were also beautifully preserved, as were insects, some with a striking iridescence still intact. Messel rocks provide a truly outstanding snapshot of life on land as the age of mammals began.
23-14
In general, the fossil record preserves some aspects of biological history well and others poorly. Fossils provide a good sense of how the forms, functions, and diversity of skeletonized animals have changed over the past 500 million years. The same is true for land plants and unicellular organisms that form mineralized skeletons. These fossils shed light on major patterns of morphological evolution and diversity change through time; their geographic distributions record the movements of continents over millions of years; and the radiations and extinctions they document show how life responds to environmental change, both gradual and catastrophic.
23.3.3 Geological data indicate the age and environmental setting of fossils.
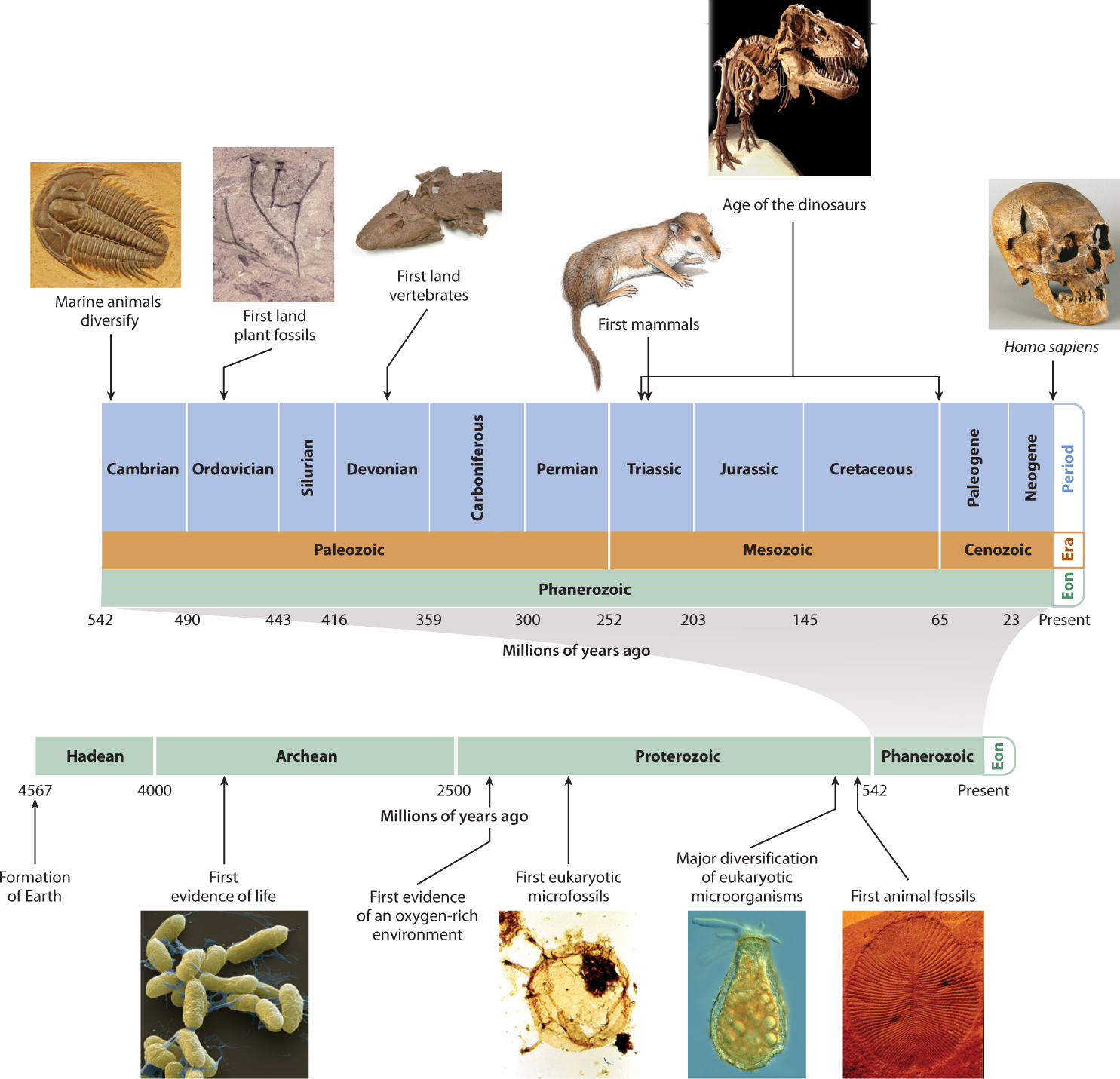
How do we know the ages of fossils? Beginning in the nineteenth century, geologists recognized that groups of fossils change systematically from the bottom of a sedimentary rock formation to its top. As more of Earth’s surface was mapped and studied, it became clear that certain fossils always occur in layers that lie beneath (and so are older than) layers that contain other species. From these patterns, geologists concluded that fossils mark time in Earth history. At first, geologists didn’t know why fossils changed from one bed to the next, but after Darwin the reason became apparent: Fossils record the evolution of life on Earth. They eventually mapped out the geologic timescale, the series of time divisions that mark Earth’s long history (Fig. 23.16).
23-15
The layers of fossils in sedimentary rocks can tell us that some rocks are older than others, but they cannot by themselves provide an absolute age. Calibration of the timescale became possible with the discovery of radioactive decay. In Chapter 2, we discussed isotopes, variants of an element that differ from one another in the number of neutrons they contain. Many isotopes are unstable and spontaneously break down to form other, more stable isotopes. In the laboratory, scientists can measure how fast unstable isotopes decay. Then, by measuring the amounts of the unstable isotope and its stable daughter inside a mineral, they can determine when the mineral formed.
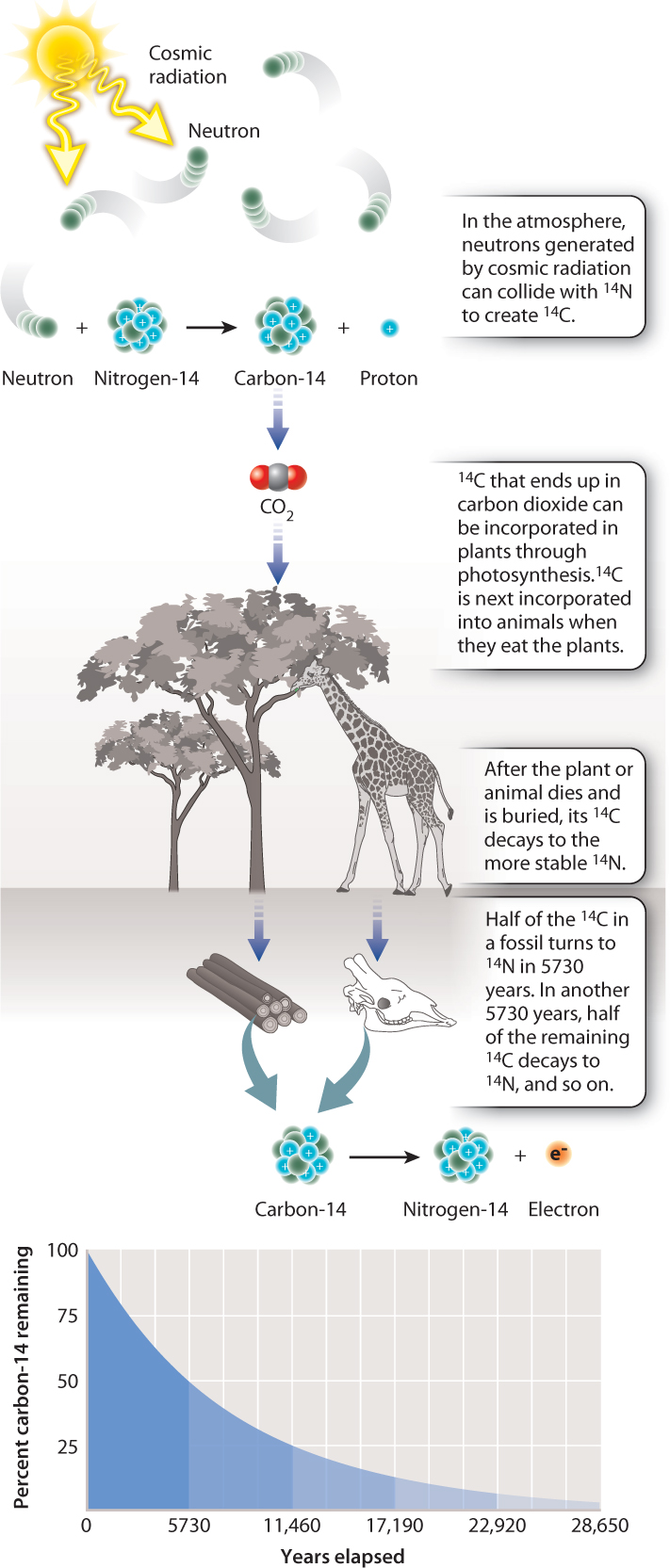
Archaeologists commonly use the radioactive decay of the isotope carbon-14, or 14C, to date wood and bone. As shown in Fig. 23.17, cosmic rays continually generate 14C in the atmosphere. Through photosynthesis, carbon dioxide that contains 14C is incorporated into wood, and animals also incorporate small amounts of 14C into their tissues when they eat plant material. After death, the unstable 14C in these materials begins to break down, losing an electron to form 14N, a stable isotope of nitrogen. Laboratory measurements indicate that half of the 14C in a given sample will decay to nitrogen in 5730 years, a period called its half-life (Fig. 23.17). Armed with this information, scientists can measure the amount of 14C in an archaeological sample and, by comparing it to the amount of 14C in a sample of known age—annual rings in trees, for example, or yearly growth coral skeletons—determine the age of the sample.
Because its half-life is so short (by geological standards), 14C is useful only in dating materials younger than 50,000 to 60,000 years. Beyond that, there is too little 14C left to measure accurately. Older geological materials are commonly dated using the radioactive decay of uranium (U) to lead (Pb): 238U breaks down to 206Pb with a half-life of 4.47 billion years, and 235U decays to 207Pb with a half-life of 704 million years. The calibration of the geologic timescale is based mostly on radiometric ages of volcanic ash beds found within sedimentary rock that contain key fossils, as well as volcanic rocks that intrude into (and so are younger than) layers of rock containing fossils. In turn, the age of fossils provide calibration points for phylogenies.
23-16
The sedimentary rocks that contain fossils also preserve, encrypted in their physical features and chemical composition, information about the environment in which they formed. Sandstone beds, for example, may have rippled surfaces, like the ripples produced by currents that we see today in the sand of a seashore or a lake margin. Enrichment in pyrite (FeS2) or other iron minerals may signal oxygen depletion.
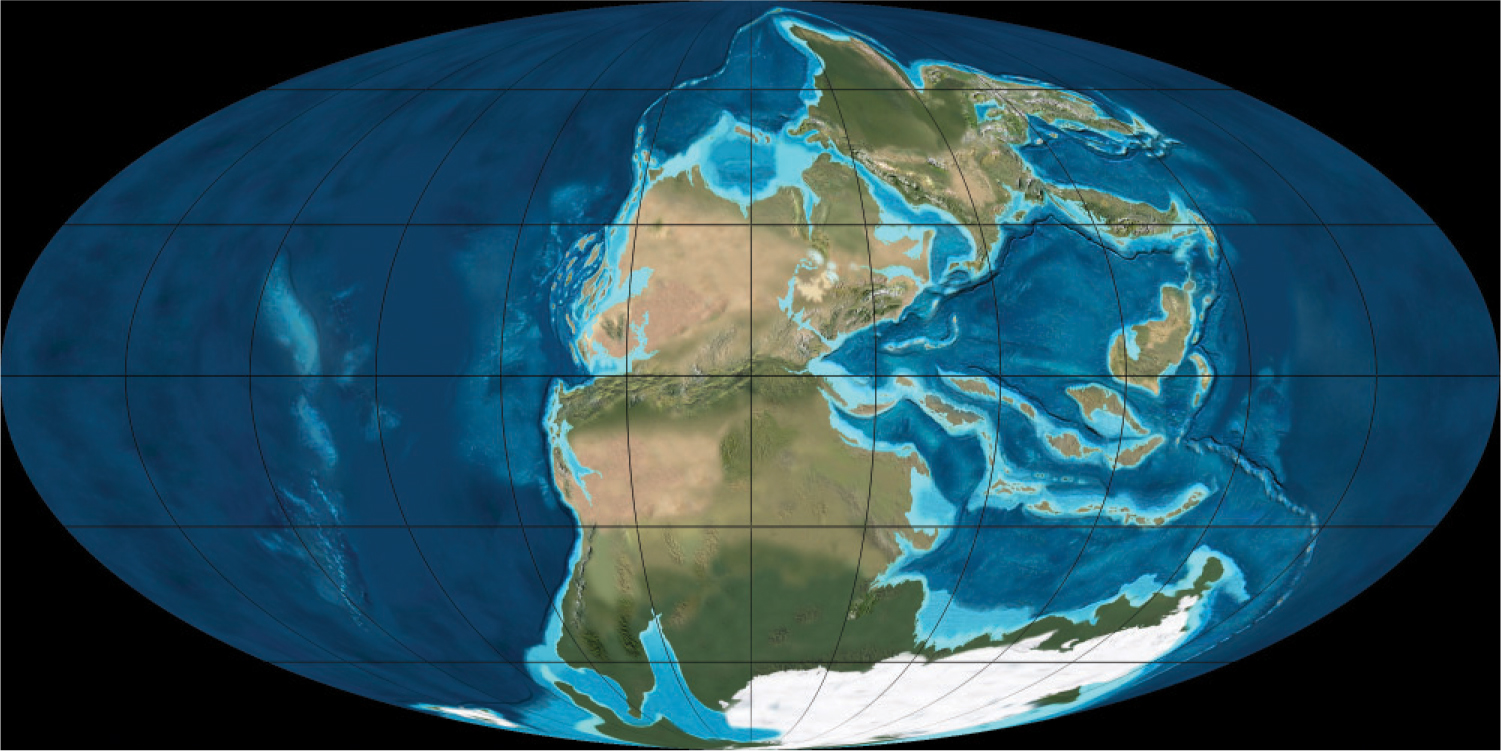
We might think our moment in geologic time is representative of the Earth as it has always existed, but nothing could be further from the truth. In the location and sizes of its continents, ocean chemistry, and atmospheric composition, the Earth we experience is unlike any previous state of the planet. Today, for example, the continents are distributed widely over the planet’s surface, but 290 million years ago they were clustered in a supercontinent called Pangaea (Fig. 23.18). Oxygen gas permeates most surface environments of Earth today, but 3 billion years ago, there was no O2 anywhere. And, just 20,000 years ago, 2 km of glacial ice stood where Boston lies today. Sedimentary rocks record the changing state of Earth’s surface over billions of years and show that life and environment have changed together through time, each influencing the other.
23.3.4 Fossils can contain unique combinations of characters.
Phylogenies hypothesize impressive morphological and physiological shifts through time—amphibians from fish, for example, or land plants from green algae. Do fossils capture a record of these transitions as they took place?
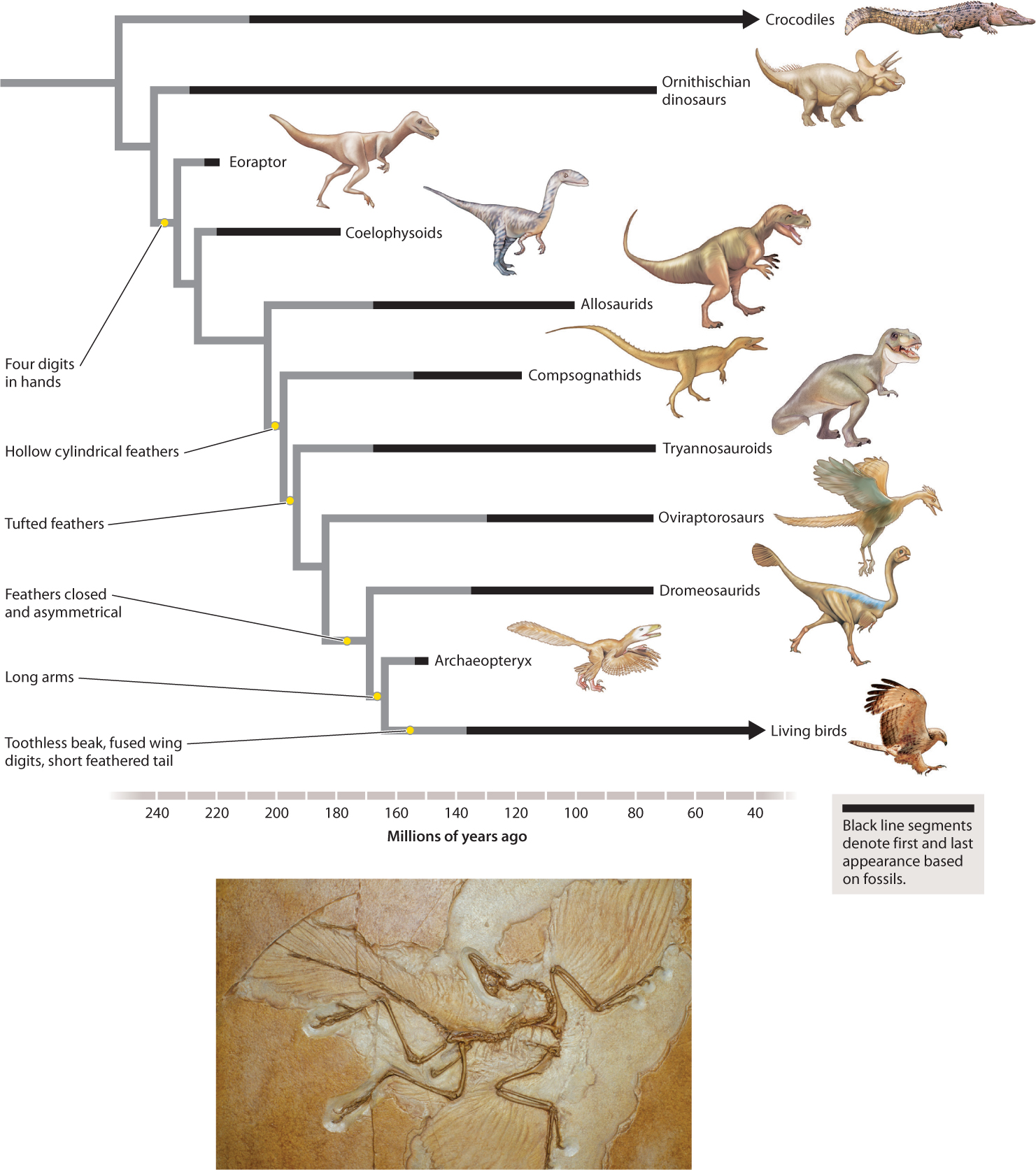
Let’s begin with an example introduced earlier in this chapter. Phylogenies based on living organisms generally place birds as the sister group to crocodiles and alligators, but birds and crocodiles are decidedly different from each other in structure—birds have wings, feathers, toothless bills, and a number of other skeletal features distinct from those of crocodiles. In 1861, just 2 years after publication of On the Origin of Species, German quarry workers discovered a remarkable fossil that remains paleontology’s most famous example of a transitional form. Archaeopteryx lithographica, now known from 11 specimens splayed for all time in fine-grained limestone, lived 150 million years ago. Its skeleton shares many characters with dromaeosaurs, a group of small, agile dinosaurs, but several features—its pelvis, its braincase, and, especially, its winglike forearms—are distinctly birdlike. Spectacularly, the fossils preserve evidence of feathers. Archaeopteryx clearly suggests a close relationship between birds and dinosaurs, and phylogenetic reconstructions that include information from fossils show that many of the characters found today in birds accumulated through time in their dinosaur ancestors. And, as noted earlier, even feathers first evolved in dinosaurs (Fig. 23.19).
Tiktaalik roseae and other skeletons in rocks deposited 375 to 362 million years ago record an earlier but equally fundamental transition: the colonization of land by vertebrates. Phylogenies show that all land vertebrates, from amphibians to mammals, are descended from fish. As seen in Fig. 23.20, Tiktaalik had fins, gills, and scales like other fish of its day, but its skull was flattened, more like that of a crocodile than a fish, and it had a functional neck and ribs that could support its body—features today found only in tetrapods. Along with other fossils, Tiktaalik captures key moments in the evolutionary transition from water to land, confirming the predictions of phylogeny.
FIG. 23.20: Can fossils bridge the evolutionary gap between fish and tetrapod vertebrates?
BACKGROUND Phylogenies based on both morphological and molecular characters indicate that fish are the closest relatives of four-legged land vertebrates.
HYPOTHESIS Land vertebrates evolved from fish by modifications of the skeleton and internal organs that made it possible for them to live on land.
OBSERVATION Fossil skeletons 390 to 360 million years old show a mix of features seen in living fish and amphibians. Older fossils have fins, fishlike heads, and gills, and younger fossils have weight-bearing legs, skulls with jaws able to grab prey, and ribs that help ventilate lungs. Paleontologists predicted that key intermediate fossils would be preserved in 380–370-million-year-old rocks. In 2004, Edward Daeschler, Neil Shubin, and Farish Jenkins discovered Tiktaalik, a remarkable fossil that has fins, scales, and gills like fish, but wrist bones and fingers, an amphibian-like skull, and a true neck (which fish lack).
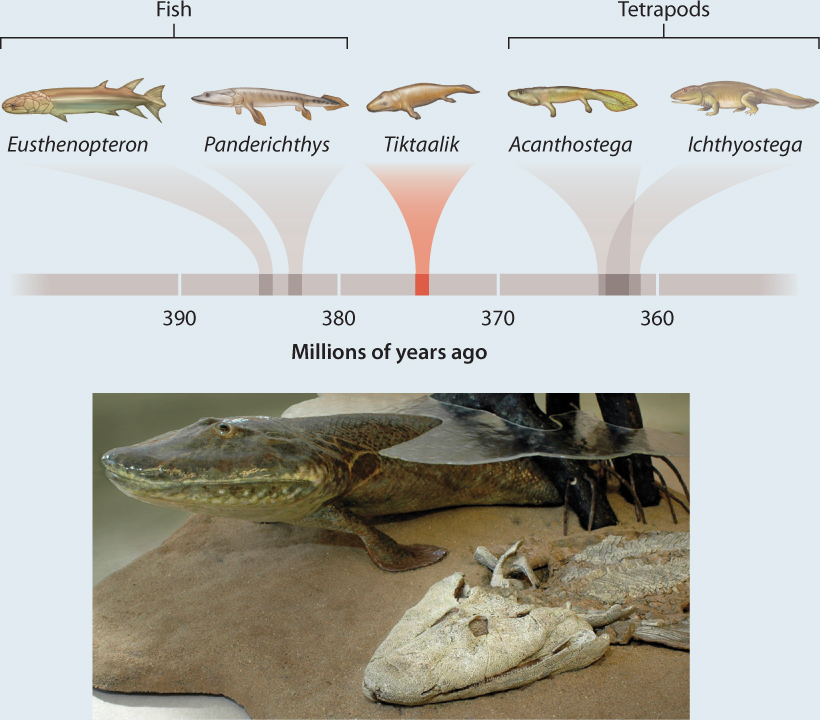
CONCLUSION Fossils confirm the phylogenetic prediction that tetrapod vertebrates evolved from fish by the developmental modification of limbs, skulls, and other features.
FOLLOW-UP WORK Research into the genetics of vertebrate development shows that the limbs of fish and amphibians are shaped by similar patterns of gene expression, providing further support for the phylogenetic connection between the two groups.
SOURCE Daeschler, E. B., N. H. Shubin, and F. A. Jenkins, Jr. 2006. “A Devonian Tetrapod-like Fish and the Evolution of the Tetrapod Body Plan.” Nature 440:757–763.
23-17
23-18
Question Quick Check 4
kzVKvUkcCvkQ0whXEmvBdC4LFUtc4/dOqLROeg0qilc2p5bRwo7lx7DJ1gV3T0KwSe4qvg/CUaQ6/ytMtUB/Ih+0FeW/tFFA/rqapoodV2Q+P0L6a7wDxVq8NUC0A7PKFSVfXcrlHgcKUHdSHT0yUlu2Gv+vlDG+GyGW01lcNqAdpZrssqN2BNTG4B1BNdFHJ4rwAuynn+vh2xAVIp6I3wqIS7nXoCm6upsl2BH4PTHpmsK223.3.5 Rare mass extinctions have altered the course of evolution.
Fossils show that life originated more than 3.5 billion years ago, but animals appeared much later, only about 600 million years ago. Fig. 23.21 graphs the number of fossil genera found for each period of the Phanerozoic (literally, the age of visible animals) Eon. Recorded diversity was low at the beginning of the Phanerozoic and, in general, has increased through time. Nonetheless, the graph clearly shows that at several moments in the past 542 million years, recorded diversity dropped catastrophically. These mass extinctions spelled the end of many previously important groups of species, but they opened up new possibilities for evolution.
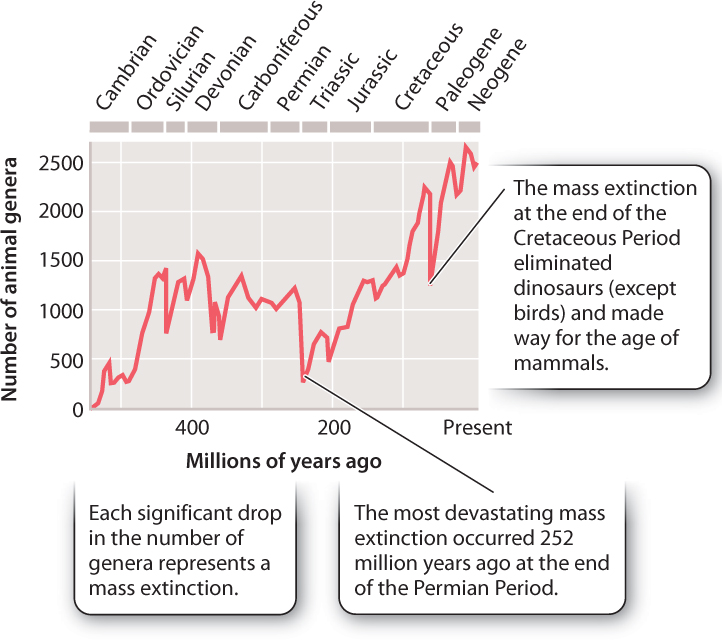
The best-known mass extinction occurred 65 million years ago, at the end of the Cretaceous Period (Fig. 23.21). On land, dinosaurs disappeared abruptly, following more than 150 million years of dominance in terrestrial ecosystems. In the oceans, ammonites, cephalopod mollusks that had long been abundant predators, also became extinct, and most skeleton-forming microorganisms in the oceans disappeared as well. As discussed in Chapter 1, a large body of geologic evidence supports the hypothesis that this biological catastrophe was caused by the impact of a giant meteorite. The mass extinction at the end of the Cretaceous removed much of the biological diversity that had built up through genetic changes over millions of years, but it had another, equally important evolutionary consequence. Survivors of the extinction could proliferate with little competition from other populations. Mammals, our own branch of the animal tree, diversified across continents once dinosaurs had disappeared from the landscape.
23-19
An even larger mass extinction took place much earlier at the end of the Permian Period, 252 million years ago (Fig. 23.21). More than 90% of all genera recorded in late Permian oceans disappeared, especially animals like corals and sponges with heavy skeletons. There is no compelling evidence for meteorite impact, so geologists have hypothesized that this mass extinction resulted from the catastrophic effects of massive volcanic eruptions.
At the end of the Permian Period, most continents were gathered into the supercontinent Pangaea (see Fig. 23.18), and so a huge ocean covered more than half the Earth. Levels of oxygen in the deep waters of this ocean fell, the result of sluggish circulation and warm seawater temperatures. Then, a massive outpouring of ash and lava erupted across what is now Siberia. Enormous emissions of carbon dioxide and methane from the volcanoes caused global warming (so even less oxygen reached deep oceans) and ocean acidification (so it was difficult for animals and algae to secrete calcium carbonate skeletons).
The three-way insult of lack of oxygen, ocean acidification, and global warming doomed many species on land and in the seas. Seascapes dominated by corals and shelled invertebrates called brachiopods disappeared. After the extinction of these organisms, new populations of mollusks, new forms of coral, and other animals we see today rose to prominence. Again, major environmental disruption removed diversity that had slowly accumulated through many millions of years but also opened up new evolutionary possibilities for the survivors.
Five great mass extinctions occurred over the past 500 million years, but only the two at the end of the Permian and Cretaceous Periods so thoroughly changed the course of evolution. At present, we live at a time of accelerating ocean acidification and global warming. Will future geologists recognize our evolutionary moment as a time of mass extinction? We discuss this possibility in Chapter 48.