24.1 THE GREAT APES
We can approach the question of our place in the tree of life in three different ways: through comparative anatomy, molecular analysis, and the fossil record. In this section, we use data from all three sources as we apply the standard methods of phylogenetic reconstruction (Chapter 23) to figure out the evolutionary relationships between humans and other mammals.
24.1.1 Comparative anatomy shows that the human lineage branches off the great apes tree.
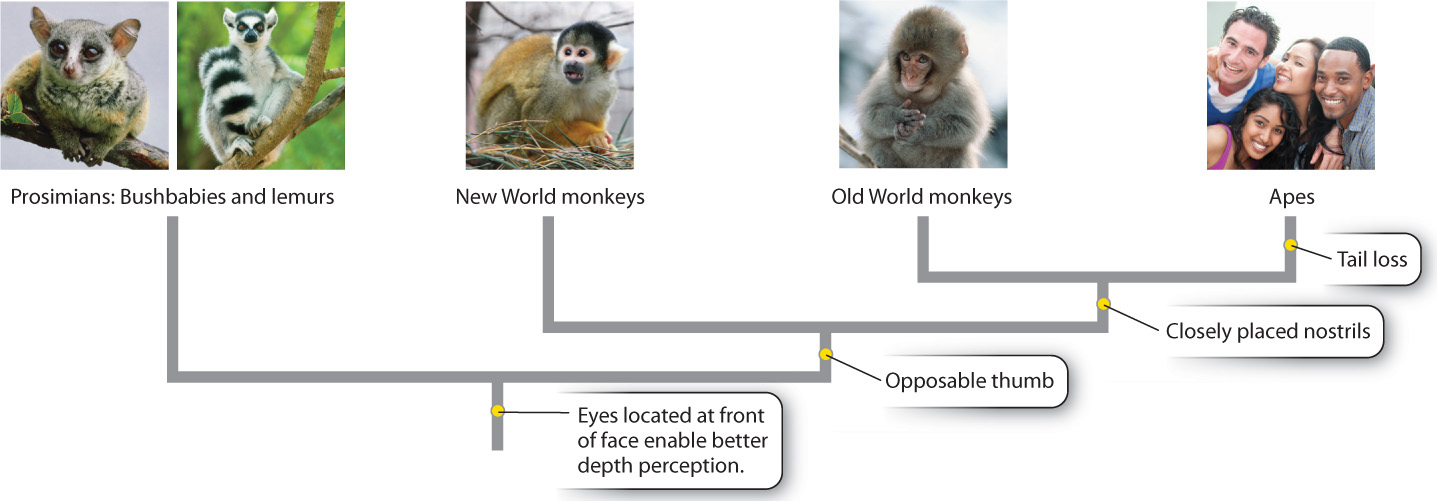
There are about 400 species of primates, which include prosimians (lemurs, bushbabies), monkeys, and apes (Fig. 24.1). All primates share a number of general features that distinguish them from other mammals, including nails rather than claws and eyes on the front of the face instead of the side, allowing stereoscopic (that is, three-dimensional) vision. Most primates also have some form of opposable thumb (a thumb that can touch the finger tips of the same hand; contrast the motion of your thumb with that of your non-opposable big toe). Prosimians are thought to represent a separate primate lineage from the one that gave rise to humans. Lemurs, which today are confined to the island of Madagascar, are thus only distantly related to humans.
24-2
Monkeys underwent independent bouts of evolutionary change in the Americas and in Africa and Eurasia, so the family tree is split along geographic lines into New and Old World monkeys. Though both groups have evolved similar habits, there are basic distinctions. There are differences between the teeth of the two groups, and in New World monkeys the nostrils tend to be widely spaced, whereas in Old World species they are closer together.
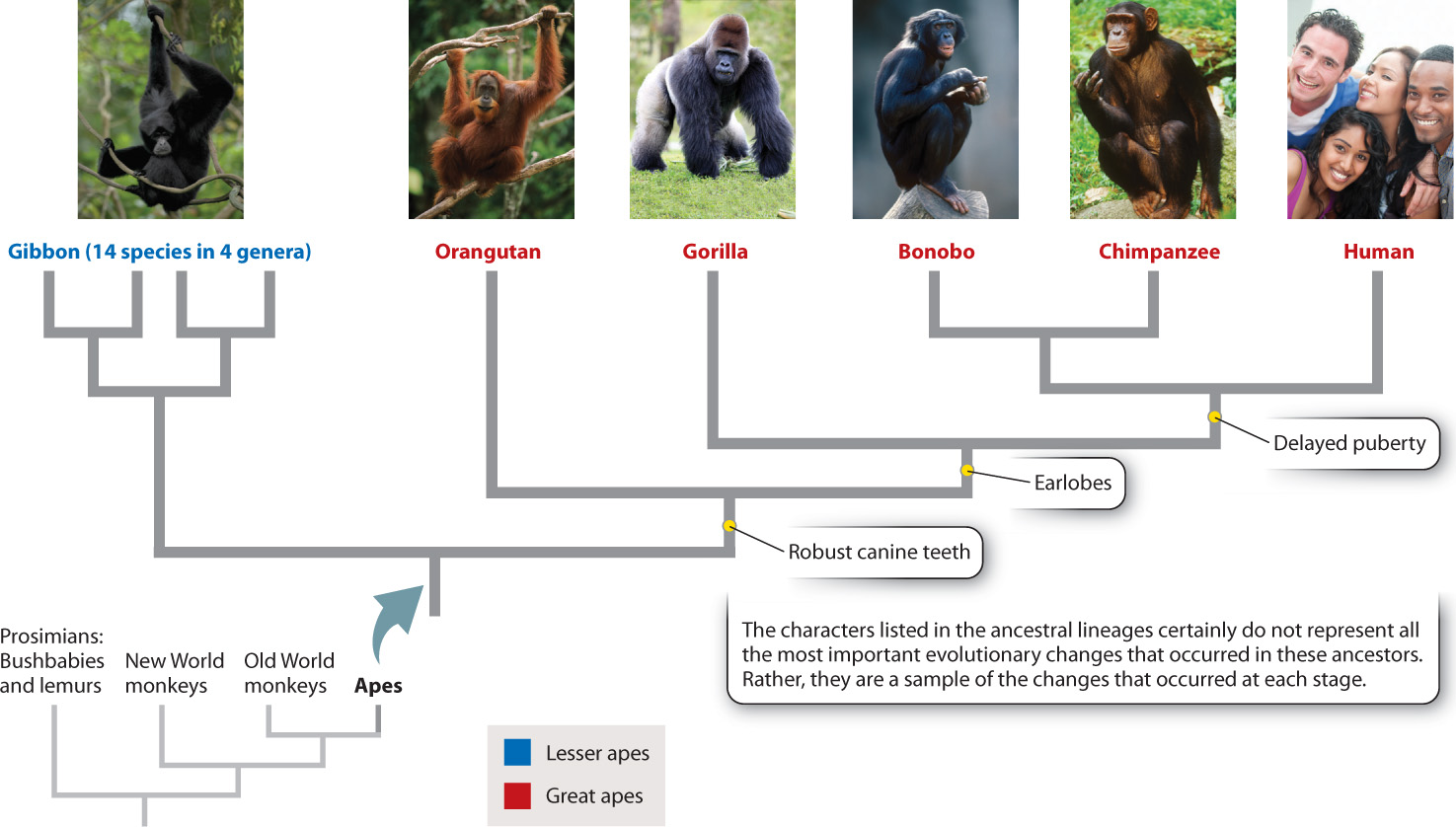
One line of Old World monkeys gave rise to the apes, which lack a tail and show more sophisticated behaviors than other monkeys. The apes are split into two groups, the lesser and the great apes (Fig. 24.2). Lesser apes include the fourteen species of gibbon, all of which are found in Southeast Asia. The great apes include the orangutan, gorilla, chimpanzee and, strictly speaking, humans. Taxonomists classify all the descendants of a specified common ancestor as belonging to a monophyletic group (Chapter 23). Thus, humans, blue whales, and hedgehogs are all mammals because all three are descended from the first mammal, the original common ancestor of all mammals. Because we trace our ancestry to the common ancestor of orangutans, gorillas, and chimpanzees, we, too, are a member of the monophyletic great ape group.
24-3
24.1.2 Molecular analysis reveals that our lineage split from the chimpanzee lineage about 5–7 million years ago.
Which great ape is most closely related to humans? That is, which is our sister group? Traditional approaches of reconstructing evolutionary history by comparing anatomical features failed to distinguish between two candidates, gorillas and chimpanzees. The implication is that humans, gorillas, and chimpanzees split into separate lineages at about the same time. It was only with the introduction of molecular methods of assessing evolutionary relationships—through the comparison of DNA and protein amino acid sequences from the different species—that we had the answer. Our closest relative is the chimpanzee (Fig. 24.2)—or, more accurately, the chimpanzees, plural, because there are two closely related chimpanzee species, the smaller of which is often called the bonobo.
Just how closely are humans and chimpanzees related? To answer this question, we need to know the timing of the evolutionary split that led along one fork to chimpanzees and along the other to us. As we saw in Chapter 21, DNA sequence differences accumulate between isolated populations or species, and they do so at a more or less constant rate. As a result, the extent of the sequence difference between two species is a good indication of the amount of time they have been separate lineages, that is, the amount of time since their last common ancestor.
The first thorough comparison of DNA molecules between humans and chimpanzees was carried out before the advent of DNA sequencing methods by Mary-Claire King and Allan Wilson at the University of California at Berkeley (Fig. 24.3). One of their methods to measure molecular differences between species relied on DNA–DNA hybridization (Chapter 12). Two complementary strands of DNA in a double helix can be separated by heating the sample. If the two strands are not perfectly complementary, as is the case if there is a basepair mismatch (for example, a G paired with a T rather than a C), less heat is required to separate the strands.
FIG. 24.3: How closely related are humans and chimpanzees?
BACKGROUND Phylogenetic analysis based on anatomical characteristics had established that chimpanzees are closely related to humans. Mary-Claire King and Allan Wilson used molecular techniques to determine how closely related the two species are.
HYPOTHESIS Despite marked anatomical and behavioral differences between the two species, the genetic distance between the two is small, implying a relatively recent common ancestor.
METHOD When two complementary strands of DNA are heated, the hydrogen bonds pairing the two helices are broken at around 95°C and the double helix denatures, or separates (Chapter 12). Two complementary strands with a few mismatches separate at a temperature slightly lower than 95°C because fewer hydrogen bonds hold the helix together. Many more mismatches between the two sequences results in an even lower denaturation temperature. Using hybrid DNA double helices with one strand contributed by each species—humans and chimpanzees, in this case—and determining their denaturation temperature, King and Wilson could infer the genetic distance (the extent of genetic divergence) between the two species.
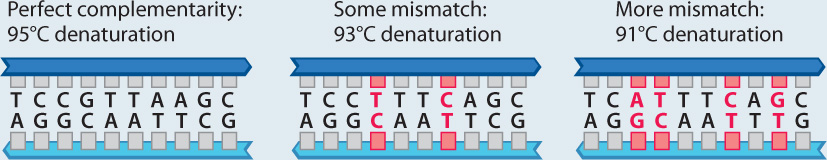
RESULTS King and Wilson found that human–chimpanzee DNA molecules separated at a temperature approximately 1°C less than the temperature at which human–human DNA molecules separate. This difference could be calibrated on the basis of studies of other species whose genetic distances were known from other methods. The DNA of humans differs from that of chimpanzees by about 1%.
CONCLUSION AND INTERPRETATION King and Wilson noted the discrepancy between the extent of genetic divergence (small) and the extent of anatomical and behavioral divergence (large) between humans and chimpanzees. They suggested that one way in which relatively little genetic change could produce extensive phenotypic change is through differences in gene regulation (Chapter 19). A small genetic change in a control region responsible for switching a gene on and off might have major consequences for the organism.
FOLLOW-UP WORK The sequences of the chimpanzee and human genomes allow us to compare the two sequences directly, and these data confirm King and Wilson’s observations.
SOURCE King, M. C., and A. C. Wilson. 1975. “Evolution at Two Levels in Humans and Chimpanzees.” Science 188:107–116.
24-4
King and Wilson used this fact to examine the differences between a human strand and the corresponding chimpanzee strand. They inferred the extent of DNA sequence divergence from the melting temperature and made a striking discovery: Human and chimpanzee DNA differ in sequence by just 1%.
King and Wilson’s conclusions have been confirmed by subsequent research. Now that we have both the human and chimpanzee genome sequences in hand, we can literally count the differences between the two sequences: The genomes differ by about 1%. The overall amount of difference between human and chimpanzee genomes goes up somewhat if you take into account segments of the genomes that are absent in one or other of the species (that is, sequences where DNA has been inserted or deleted in one lineage or the other).
Because the amount of sequence difference is correlated with the length of time the two species have been isolated, we can convert the divergence results into an estimate of the timing of the split between the human and chimpanzee lineages. That split occurred about 5–7 million years ago. All the extraordinary characteristics that set our species apart from the rest of the natural world—those attributes that are ours and ours alone—arose in just 5–7 million years.
Question Quick Check 1
BGgXyDfV88w8VTOuUv6yuNRnQ5up9QH/F2SvTRR4MNZPwu+Pc7cvUyGTrBXp/iDe24.1.3 The fossil record gives us direct information about our evolutionary history.

Molecular analysis is a powerful tool for comparing species and populations within species. It allows us to compare humans and chimpanzees and look at differences among groups of humans or groups of chimpanzees. But we are limited in what we can study because we need samples from living individuals. We can study the end products of evolution but not the intermediate stages, which have long since disappeared and cannot provide us with DNA samples. For a full picture of human evolution, we must turn to fossils (Chapter 23).
For the first several million years of human evolution, all the fossils from the human lineage are found in Africa. This is not surprising. Charles Darwin himself noted that it was likely that the human lineage originated there, as humans’ two closest relatives, chimpanzees and gorillas, live only in Africa. The fossil material varies in quality, and a great deal of ingenuity is often required to reconstruct the appearance and attributes of an individual from fragmentary fossil material. It’s hard to determine whether two fossil specimens with slight differences belong to the same or different species. And, of course, interbreeding—the criterion that defines a species (Chapter 22)—cannot be applied to fossils. As a result, experts disagree over the details of the human fossil record. However, several robust general conclusions may be drawn.
The members of all the different species in the lineage leading to humans are called hominins. Let’s start with the earliest known member of the hominin lineage, Sahelanthropus tchadensis (Fig. 24.4). Discovered in Chad in 2002, the skull of S. tchadensis combines both modern (human) and ancestral features. S. tchadensis, which has been dated to about 7 million years ago and has a chimpanzee-sized brain but hominin-type brow ridges, probably lived shortly after the split between the hominin and chimpanzee lineages, although this conclusion is controversial.
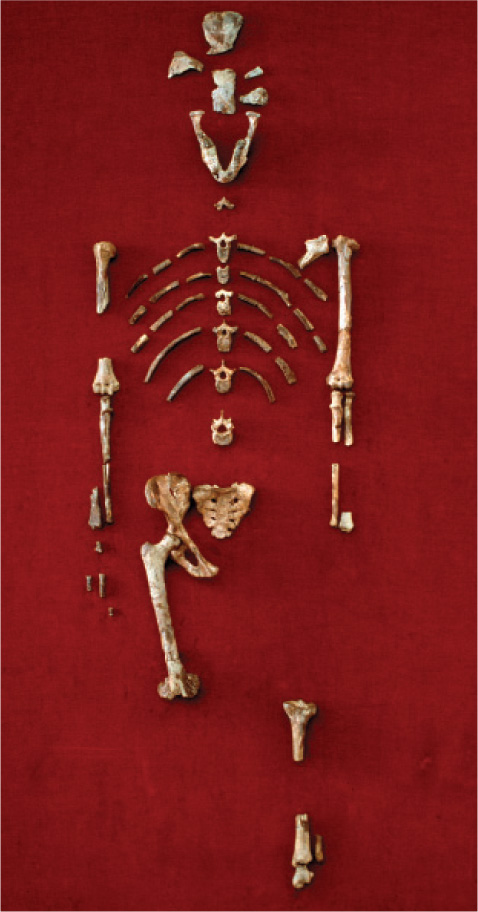
An important early hominin, dating from about 4.4 million years ago, is a specimen of Ardipithecus ramidus from Ethiopia. This individual, known as Ardi, was capable of walking upright, using two legs on the ground but all four limbs in the trees. An unusually complete early hominin fossil, Lucy, found in 1974 at Hadar, Ethiopia, represents the next step in the evolution of hominin gait: She was fully bipedal, habitually walking upright (Fig. 24.5). Her name comes from the Beatles’ song “Lucy in the Sky with Diamonds,” which was playing in the paleontologists’ field camp when she was unearthed. This fossil dates from around 3.2 million years ago. Lucy was a member of the species Australopithecus afarensis and was much smaller than modern humans, less than 4 feet tall, and had a considerably smaller brain (even when the difference in body size is taken into consideration). In many ways, then, Lucy was similar to the common ancestor of the human and chimpanzee lineages, except that she was bipedal.
24-5
Remarkably, the hominin lineage produced many different species in Africa, so that there were at times as many as three species living at the same time. Fig. 24.6 shows the main hominin species and the longevity of each in the fossil record, along with suggested evolutionary relationships among the species. Note that all hominins have a common ancestor, but not all of these lineages lead to modern humans, producing instead other branches of the hominin tree that ultimately went extinct.
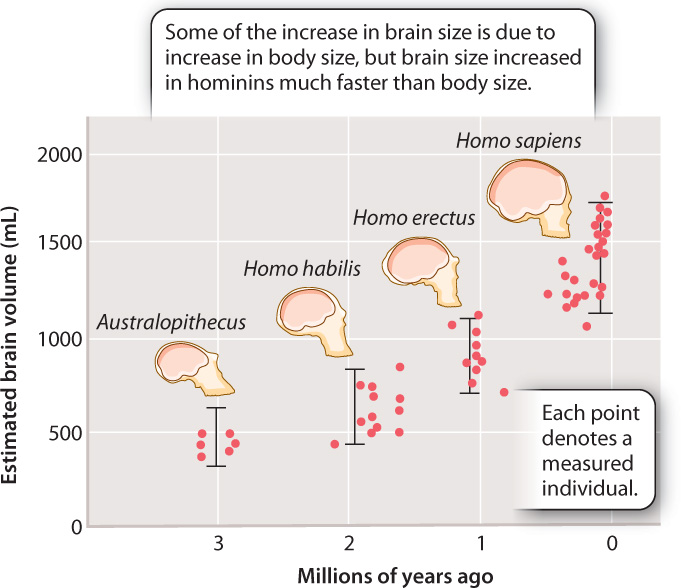
A number of trends can be seen when we look over the entire record. Body size increased, and most striking is the increase in size of the cranium and therefore, by inference, of the brain, as shown in Fig. 24.7.
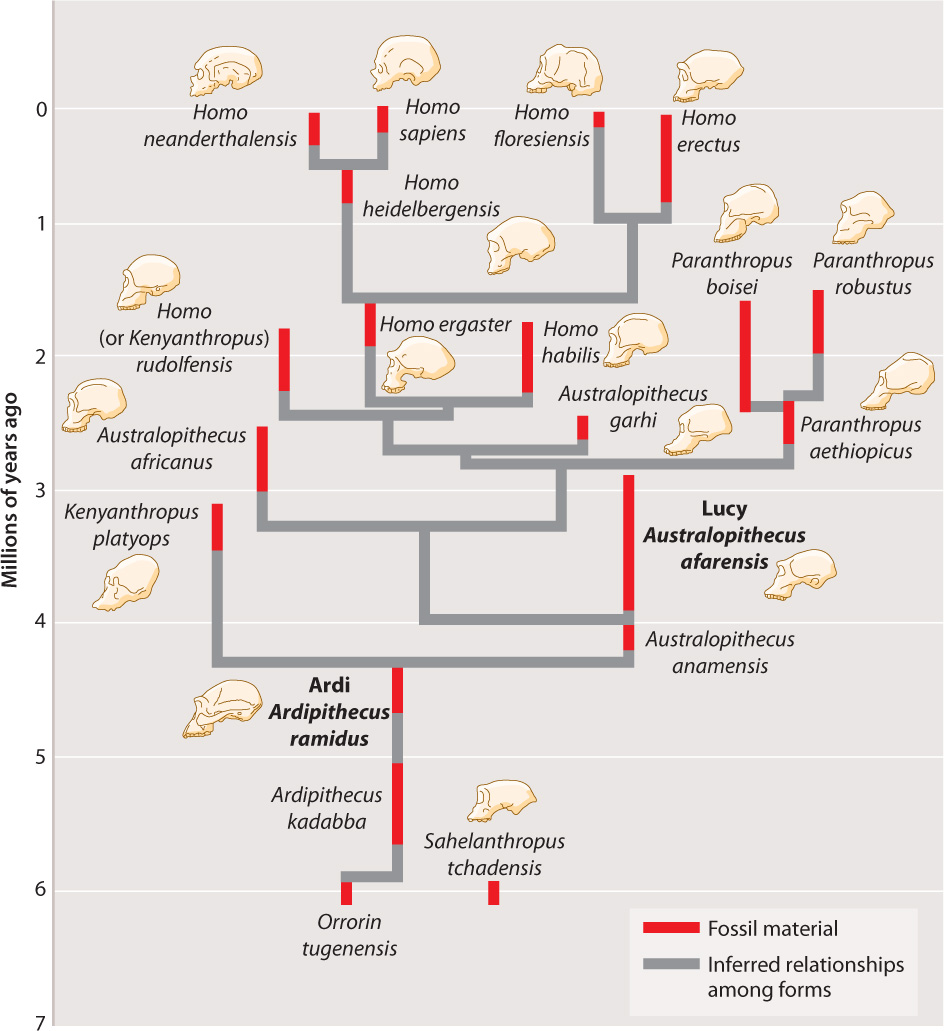
The fossil record indicates that about 2 million years ago, the hominin lineage ventured out of Africa. The first hominin that left Africa is sometimes called Homo ergaster and sometimes Homo erectus. The confusion stems from the controversies that surround the naming of fossil species. Some researchers contend that H. ergaster is merely an early form of H. erectus. We designate this first hominin Homo ergaster and a later, descendant species, H. erectus (see Fig. 24.6). The naming details, however, are relatively unimportant. What matters is that some hominins first left Africa about 2 million years ago. Fossils have been found throughout Eurasia, though not in Australia or the Americas. It was from relatives of Homo ergaster that our species, Homo sapiens, derived.
Another species closely related to Homo sapiens was Homo neanderthalensis, whose fossils appear in Europe and the Middle East. Thicker boned than us, and with flatter heads that contained brains about the same size as, or slightly larger than, ours, Neanderthals first appeared in the fossil record around 600,000 years ago and disappeared around 30,000 years ago. As we will see, genetic analysis suggests that this disappearance was perhaps not as complete as the fossil record suggests.
Another species that probably ultimately derived from the original H. ergaster emigration from Africa became extinct only about 12,000 years ago. This was H. floresiensis, known popularly as the Hobbit. H. floresiensis is peculiar: Limited to the Indonesian island of Flores, adults were only just over 3 feet tall. Some have suggested that H. floresiensis is not a genuinely distinct species, but, rather, is an aberrant H. sapiens. Plenty of evidence, however, suggests that H. floresiensis is a distinct species derived from an archaic Homo species, probably H. erectus. Mammals often evolve small body size on islands because of the limited availability of food.
24-6