25.3 THE CARBON CYCLE, ECOLOGY, AND EVOLUTION
As you sit quietly by a woodland pond, the carbon cycle is at work all around you. In the forest, plants generate carbohydrates as they photosynthesize, and algae and photosynthetic bacteria are doing the same in the pond. These organisms are primary producers, generating organic compounds that will provide food for other organisms in the local environment.
An insect grazes on leaves, and ducks feed on both insects and algae. Ducks and insects obtain the carbon they need for growth and reproduction from the foods they eat, and they also gain energy by respiring food molecules. Such organisms are consumers, and they include all the animals in both pond and forest. Beneath our feet, similar interactions play out on a microscopic scale, especially among the fungi, protists, and bacteria that recycle the organic carbon in soil back to CO2.
The complementary metabolic processes of photosynthesis and respiration cycle carbon through forest and pond communities. Furthermore, as we will see in the next chapter, complementary metabolic processes also cycle nitrogen, sulfur, and other elements required for life. By continually recycling materials, biogeochemical cycles, which involve both biological and physical processes, sustain life over long intervals. In their absence, life could hardly have persisted for 4 billion years.
25.3.1 Food webs trace the movement of carbon through communities.
In biological communities across the planet, carbon and energy are packaged by photosynthetic organisms and transferred to heterotrophs (Chapter 6) that graze on plants or photosynthetic microorganisms. Carnivorous organisms may eat these grazers, and they, in turn, may provide food for still other carnivores. Eventually, having passed from one consumer to another, the carbon originally fixed by photosynthesis is returned to the atmosphere by the respiration of fungi and other decomposers that break down dead tissues.
The linear transfer of carbon from one organism to another is called a food chain. Because most heterotrophs within a community can consume or be consumed by a number of other species, biologists often prefer to speak of food webs, a term that provides a better sense of the complexity of interactions within the carbon cycle (Fig. 25.13). Food webs define the interactions among organisms in ponds, forests, and many other habitats. Simply put, the carbon cycle underpins the ecological structure of biological communities (Chapter 47).
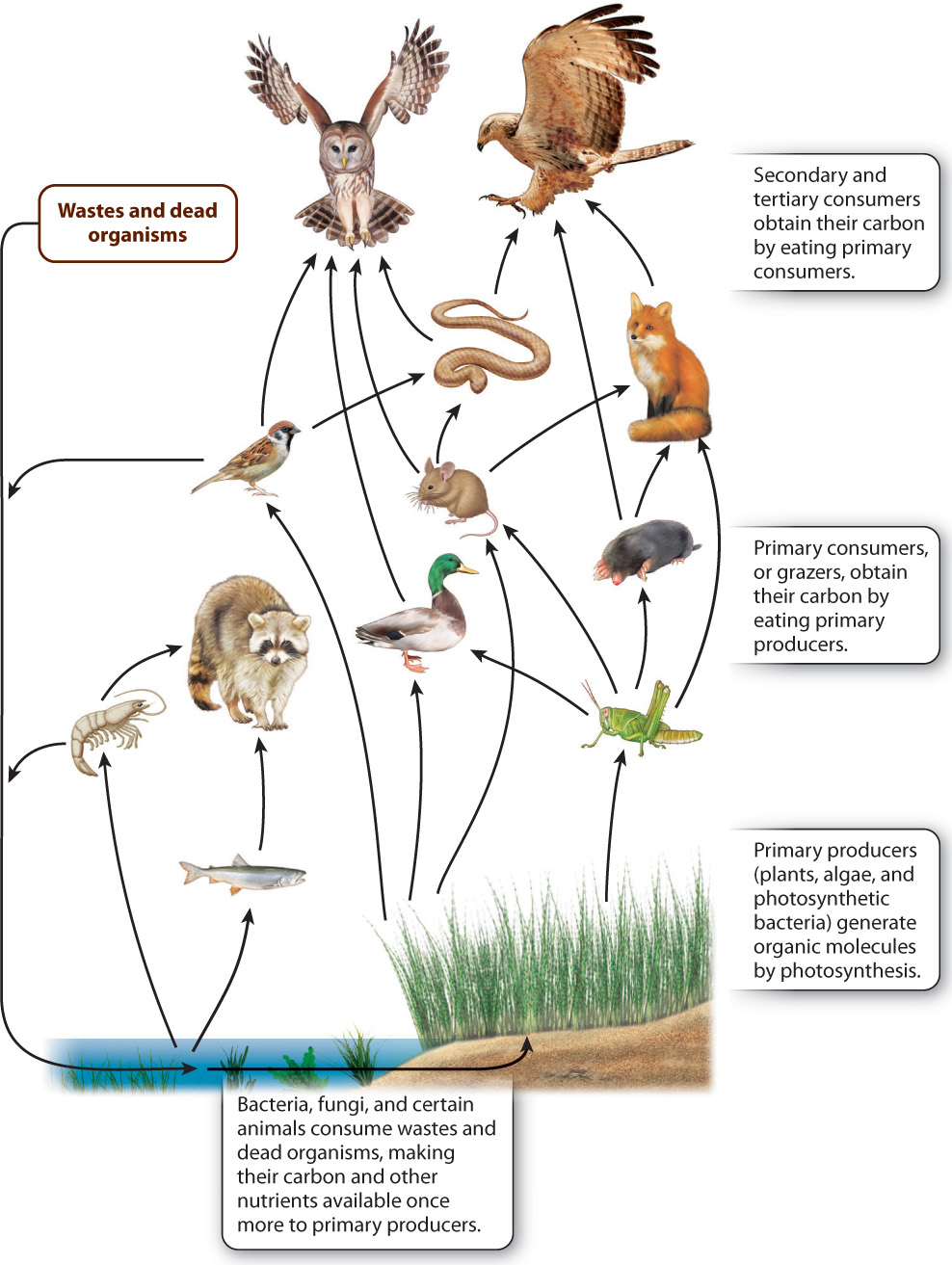
25-13
25.3.2 Trophic pyramids trace the flow of energy through communities.
Throughout this chapter and earlier ones, we have emphasized that photosynthesis and respiration transfer energy as they cycle carbon. Carbon is removed from the environment by photosynthetic organisms and passed from one consumer to another, eventually returning to the environment by respiration carried out by consumers and decomposers. At the same time as they remove carbon from the atmosphere, photosynthetic organisms also harvest energy from sunlight and convert it into chemical energy that is stored in the bonds of organic molecules. Consumers store some of the energy in their food as fat, starch, or other organic molecules, but they use much of their food energy to do work or generate heat. The energy used for work or dissipated as heat cannot be captured again by photosynthetic organisms. Whereas carbon cycles through communities, energy transfer is directional.
Within a biological community, the various producers and consumers are commonly grouped into functionally similar sets called trophic groups, from trophé, the Greek word for “nourishment.” Diagrams showing the amount of energy available at each level to feed the next are called trophic pyramids (Fig. 25.14). These diagrams have a pyramidal shape because the biomass of primary producers generally much larger than the biomass of primary consumers, and the biomass of primary consumers is, in turn, much greater than that of secondary consumers, and so on. The reason for this is that energy transfer from level to level is inefficient. Because of wastes, work, and heat dissipation, only about 10% to 15% of the energy available in biomass at one level gets incorporated into biomass at the next level. Like food webs, trophic pyramids relate community structure to the fundamental biological processes at work in the carbon cycle.
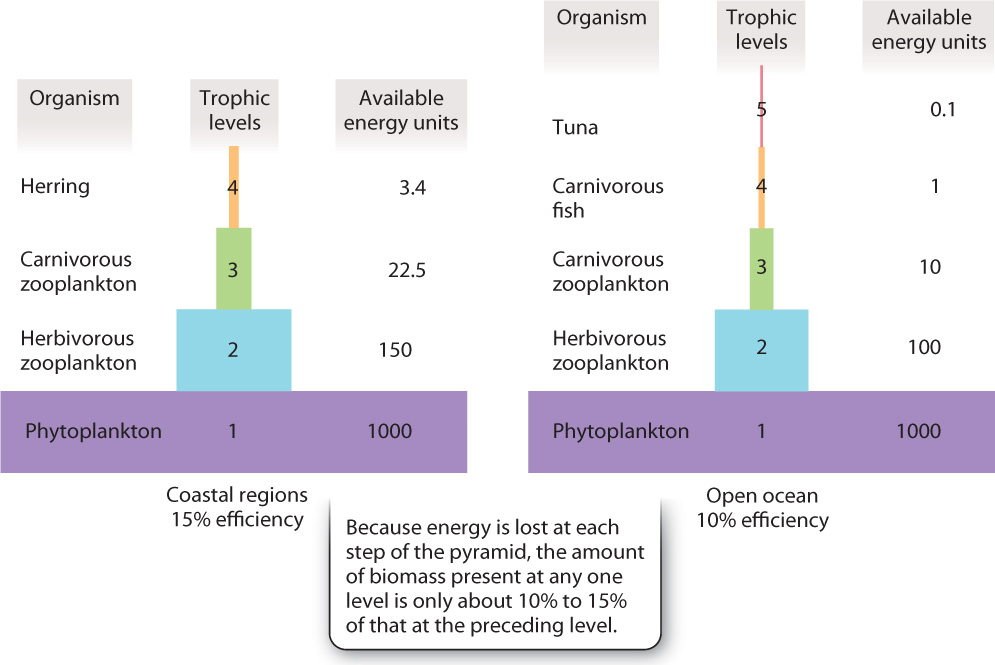
Question Quick Check 3
+c5rtSUU44kQk3NH1bw7yyJaVJRMSOPVDJagRFiLQicOoiOZk6dO4/DzYxUBPhlew8pi0pfObLYpS1UNLJdoGj7PAmDcYSZG25.3.3 The diversity of photosynthetic organisms reflects adaptation to a wide range of environments.
An estimated 500,000 species of photosynthetic organisms fuel the carbon cycle in all but a few deep-sea and subterranean environments. In terms of carbon and energy metabolism, all these species do pretty much the same thing. Why, then, is there such a diversity of photosynthetic organisms? Why don’t just a few species dominate the world’s photosynthesis?
The example of the pond and forest helps to make the basis of photosynthetic diversity clear. In the forest community, the leaves of several different tree species form a photosynthetic canopy above the forest floor. Below, shrubs grow, making use of light not absorbed by the leaves above them. And below the shrubs are grasses, herbs, ferns, and mosses that can grow in the reduced light levels of the forest floor. In the adjoining pond, a few species of aquatic plants line the water’s edge, but beyond that algae and photosynthetic bacteria dominate photosynthesis, with some species anchored to the pond bottom and others floating in the water column.
On the largest scale, then, the reason for photosynthetic diversity is obvious—cyanobacteria, algae, and land plants all perform comparable metabolic functions in different habitats. This kind of specialization for different environments also applies on a finer scale, although it is complicated by grazing and environmental disturbances such as fire and landslides (Chapter 47). In the forests of New England, the tree species that thrive in wet bottomlands differ from those found on well-drained hillsides. And seasonally dry woodlands in southern California support yet another set of plant species. In general, plants of varying size, shape, and physiology inhabit physically and biologically distinct environments, and the same is true of photosynthetic organisms in water.
Thus, the immense diversity of photosynthetic organisms found today reflects not so much evolutionary variations in the biochemistry of photosynthesis (although some of that occurs; see Chapter 29) as structural and physiological adaptations. These adaptations allow the effective gathering of light, nutrients, and—critical to life on land—water, in widely varying local environments. Natural selection, acting on local populations, links the diversity of photosynthetic organisms to the carbon cycle.
25-14
25.3.4 The diversity of respiring organisms reflects many sources of food.
About 10 million species help to cycle carbon through respiration. These include the plants, algae, and bacteria that generate carbohydrates by photosynthesis, as well as legions of animals, fungi, and microorganisms that obtain both carbon and energy from preexisting organic compounds.
Heterotrophic bacteria, amoebas, and humans may use essentially the same biochemical pathway to respire organic molecules, but they differ markedly in how they feed and, therefore, what they can eat. Bacteria (and also fungi) absorb molecules from their environment, but amoebas and many other eukaryotic microorganisms can capture and ingest cells—they are capable of predation. Animals capture prey, as well, but commonly feed on organisms far too large for an amoeba to eat. As photosynthetic organisms have adapted structurally and physiologically to local environments across the globe, heterotrophs have adapted by means of locomotion, mouth and limb specialization, perception, and behavior to obtain their food.
25.3.5 The carbon cycle provides a framework for understanding life’s evolutionary history.
The world did not always support the biological diversity we see today. Indeed, 2 billion years ago, the carbon cycle was anchored by photosynthetic bacteria and microbial heterotrophs because there were no plants or animals (Fig. 25.15). Subsequently, algae gained an ecological foothold as nutrients such as nitrate became more widely available, and single-celled eukaryotic heterotrophs expanded their reach by gathering food in ways not possible for bacteria—that is, by ingesting cells and other types of particulate food. With the evolution of multicellularity, animals and seaweeds added further complexity to carbon cycling in the sea. Eventually, some aquatic green algae evolved the capacity to live on land, and animals soon followed, building unprecedented levels of diversity on our planet.

The history of life is one of accumulating variety and complexity, based mostly on the great biological components of the carbon cycle: photosynthesis and respiration. Indeed, the carbon cycle did more than provide a framework for accumulating diversity; it changed the very nature of Earth surface environments in ways that enabled new types of organism to evolve.
25-15
When life began, the atmosphere and oceans contained little or no oxygen gas. How did our present environment come to be? The key lies in the formulas for photosynthesis and respiration, shown in Fig. 25.2. Photosynthesis and respiration not only cycle carbon, they cycle oxygen (and water) as well. When photosynthesis and respiration are in balance, the oxygen generated by photosynthesis is completely consumed by respiration. But when some of the organic carbon generated by photosynthesis is buried in sediments, oxygen production can exceed oxygen consumption. In other words, because sedimentary organic matter burial can break the tight coupling depicted in Fig. 25.2, it can facilitate an increase in O2 levels that may accumulate in the atmosphere and oceans. The geologic history of oxygen shown in Fig. 25.16 results from the interactions through time between the carbon (and sulfur) cycle with plate tectonics. Our oxygen-rich world is the result.
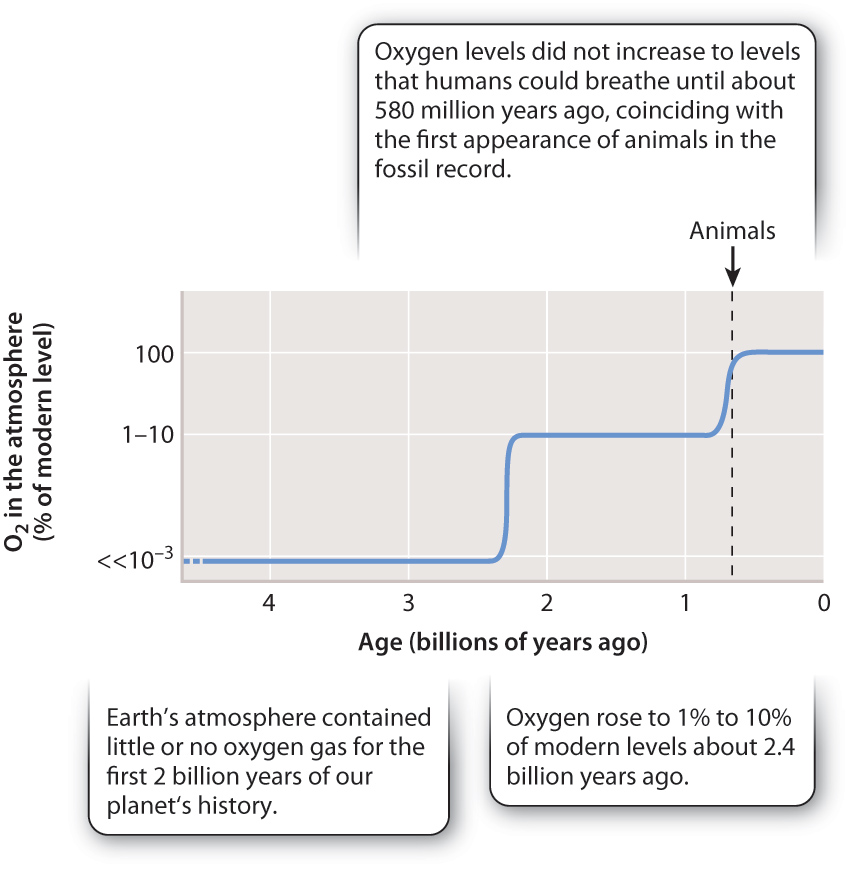
In the chapters that follow, we examine life’s diversity in terms of function, phylogeny, and ecology, but don’t be fooled into thinking that these are discrete topics: They are inextricably intertwined. The remarkable diversity of life on Earth reflects the ecology and evolution of populations that interact to cycle carbon through the biosphere.