26.1 TWO PROKARYOTIC DOMAINS
Chapter 5 outlined the two distinct ways that cells are organized internally. Eukaryotic cells, like those that make up our bodies, have a membrane-bound nucleus and organelles that form separate compartments for many cell functions. Prokaryotic cells have a simpler organization. No membrane surrounds the prokaryotic cell’s DNA, and there is little in the way of cell compartments. Prokaryotic cell organization is an ancestral character for life as a whole—that is, it is a feature that was present in the last common ancestor of all organisms alive today. The group defined traditionally as the prokaryotes, however, is paraphyletic in that it excludes some descendants of the last common ancestor of all living things, namely eukaryotic organisms (Chapter 23).
Bacteria and Archaea are the two great domains characterized by prokaryotic cell structure. These organisms are present almost everywhere on Earth’s surface. What they lack in complexity of cell structure, these tiny cells more than make up for in their dazzling metabolic diversity. As will become clear over the course of this chapter, Bacteria and Archaea underpin the efficient operation of ecosystems on our planet.
26.1.1 The bacterial cell is small but powerful.
Because of their small size and deceptively simple cell organization, bacteria were long dismissed as primitive organisms, distinguished mostly by the eukaryotic features they lack: They have no membrane-bounded nuclei, no energy-producing organelles, no sex. This point of view turns out to be more than a little misleading. Bacteria are the diverse and remarkably successful products of nearly 4 billion years of evolution. On present-day Earth, bacterial cells outnumber eukaryotic cells by several orders of magnitude.
26-2
Fig. 26.1 illustrates the bacterial cell, which was briefly introduced in Chapter 5. The cell’s DNA is present in a single circular chromosome, unlike the multiple linear chromosomes characteristic of eukaryotic cells. Many bacteria carry additional DNA in the form of plasmids, small circles of DNA that replicate independently of the cell’s circular chromosome. In general, plasmid DNA is not essential for the cell’s survival, but it may contain genes that have adaptive value under specific environmental conditions. No nuclear membrane separates DNA from the surrounding cytoplasm, and so transcribed mRNA is immediately translated into proteins by ribosomes.
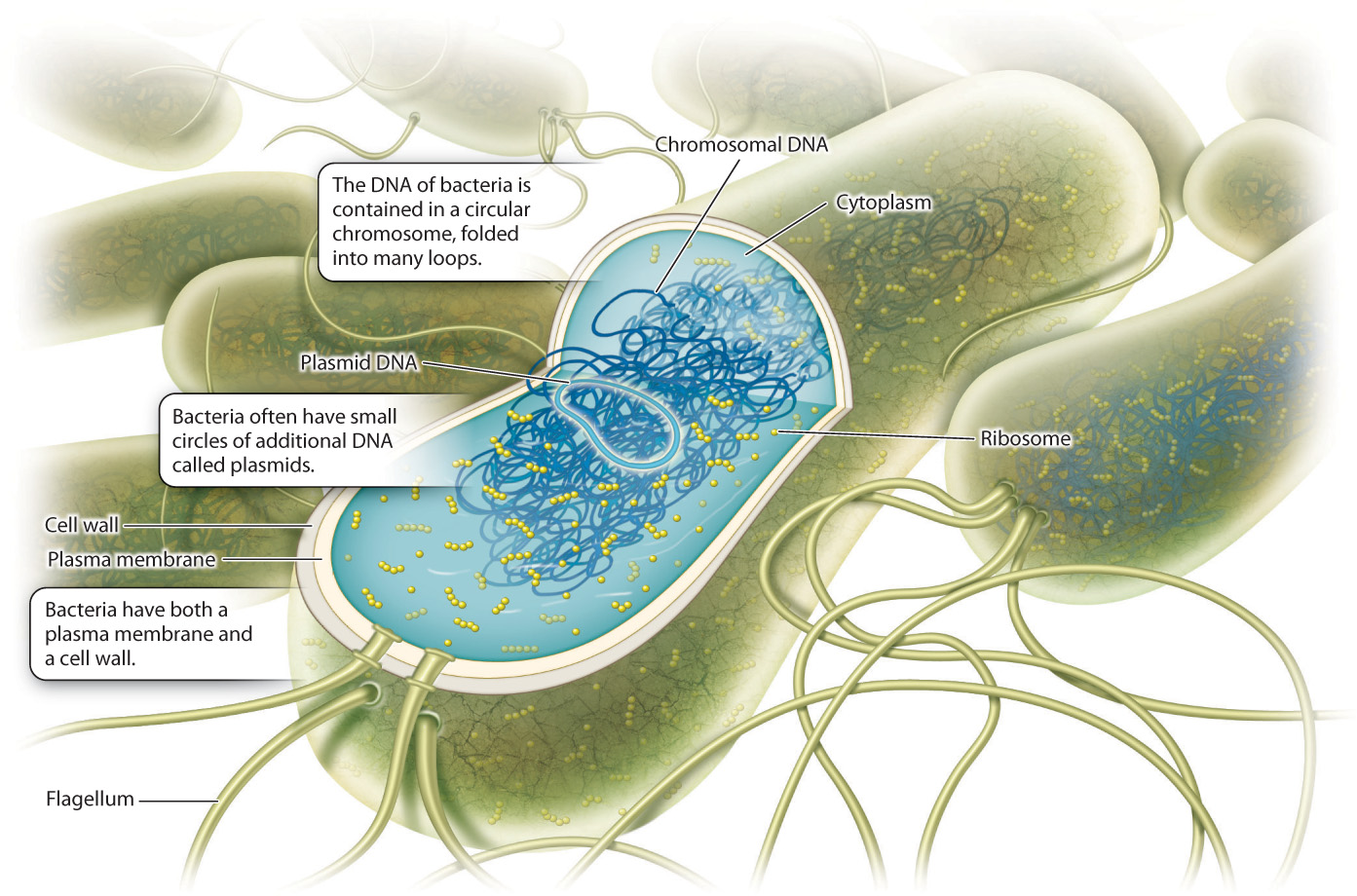
Bacteria lack the membrane-bounded organelles found in eukaryotic cells. Instead, cell processes such as metabolism are carried out by proteins that float freely in the cytoplasm or are embedded in the plasma membrane. A few bacteria, notably the photosynthetic bacteria, contain internal membranes. The light reactions of photosynthetic bacteria take place in association with membranes distributed within the cytoplasm.
Structural support is provided by a cell wall made of peptidoglycan, a complex polymer of sugars and amino acids (Chapter 10). Some bacteria have thick walls made up of multiple peptidoglycan layers, while others have thin walls surrounded by an outer layer of lipids. For many years, it was believed that bacteria lacked the cytoskeletal framework that organizes cytoplasm in eukaryotic cells. However, careful studies now show that bacteria do have an internal scaffolding of proteins that plays an important role in determining the shape, polarity, and other spatial properties of bacterial cells.
26.1.2 Diffusion limits cell size in bacteria.
Most bacterial cells are tiny: The smallest are only 200–300 nanometers (nm) in diameter, and relatively few are more than 1–2 micrometers (μm) long. Why are bacteria so small? The answer has to do with diffusion, the random motion of molecules, a critical process introduced in Chapter 5. When there is a concentration difference, diffusion results in net movement of molecules from a region of higher concentration to a region of lower concentration. If you could watch the movement of any particular molecule, you would see it is random, sometimes moving in one direction and sometimes in another. On average, however, more molecules move from a region with a higher concentration of the molecule to a region with a lower concentration of the molecule than in the other direction. Net movement stops only when the two regions achieve equal concentration, but diffusion continues. Photosynthetic bacteria gain the carbon dioxide they need by the diffusion of CO2 from the environment into the cell, and that is also how respiring bacteria take in small organic molecules and oxygen.
26-3
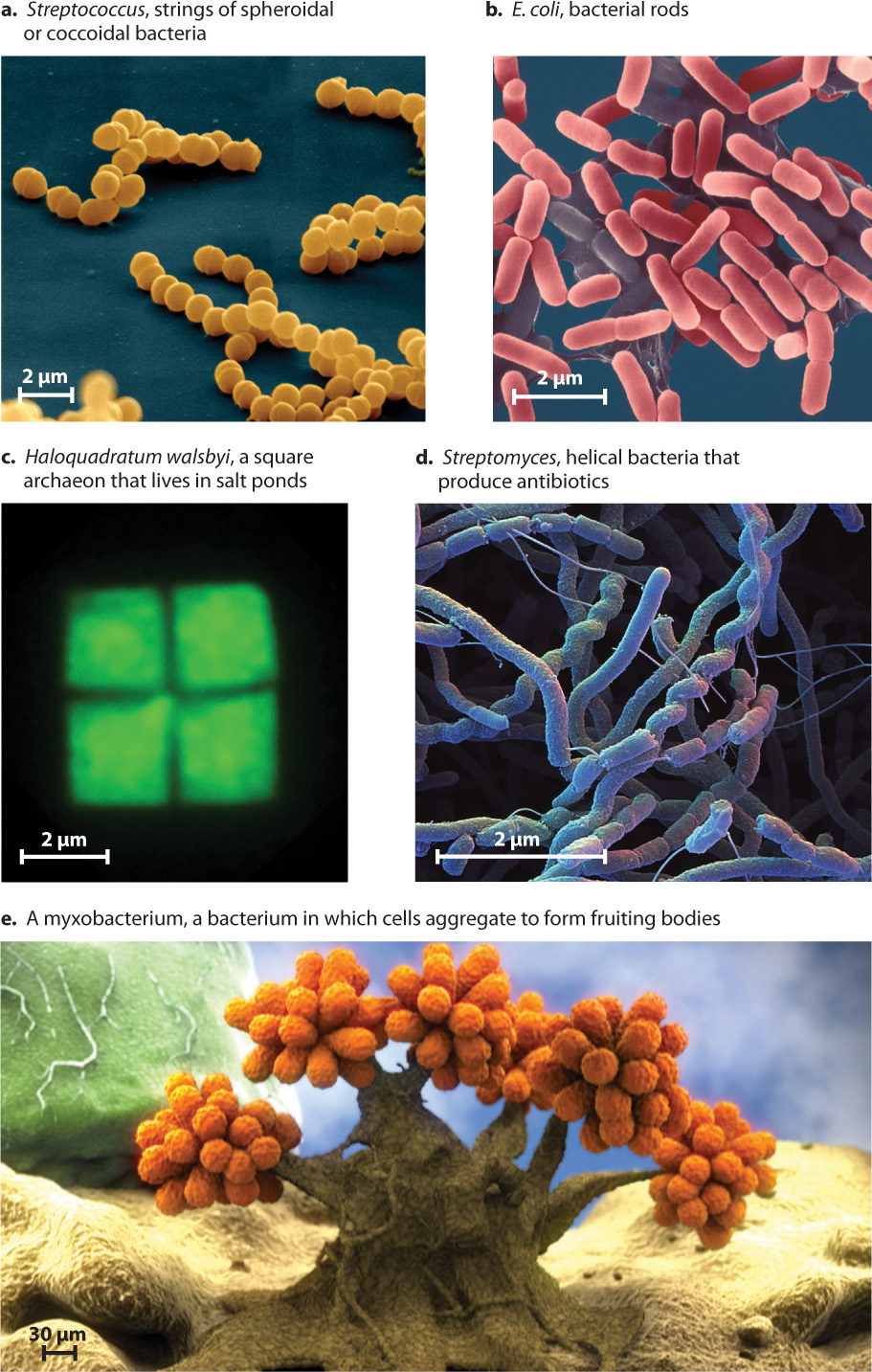
Diffusion explains why bacterial cells tend to be small. A small cell has more surface area in proportion to its volume, and so the interior parts of a small cell are closer to the surrounding environment than those of a larger cell. As a consequence, slowly diffusing molecules do not have to travel far to reach every part of a small cell’s interior. The surface area of a spherical cell—the area available for taking up molecules from the environment—increases as the square of the radius. However, the cell’s volume—the amount of cytoplasm that is supported by diffusion—increases as the cube of the radius. Therefore, as cell size increases, it becomes harder to supply the cell with the materials needed for growth. For this reason, most bacterial cells are small spheres, rods, spirals, or filaments that facilitate diffusion of molecules into cell interiors (Fig. 26.2).
A few exceptional bacteria exceed 100 μm in maximum dimension. For example, Thiomargarita namibiensis, a bacterium that lives in oxygen-poor sediments off the coast of southwestern Africa, has a total volume about 100,000,000 times larger than that of Escherichia coli (Fig. 26.3). But, in one sense, T. namibiensis cheats: 98% of its volume is taken up by a large vacuole, so the metabolically active cytoplasm is restricted to a thin film around the cell’s periphery. Thus, the distance through which nutrients move by diffusion is only a few miocrometers, as in many other bacteria.
Some bacteria are multicellular, forming simple filaments or sheets of cells. More unusual are myxobacteria, which aggregate to form multicellular reproductive structures that are composed of several distinct cell types (see Fig. 26.2e).
26-4
26.1.3 Horizontal gene transfer promotes genetic diversity in bacteria.
Bacterial genomes are generally smaller than those of eukaryotes, in part because bacteria lack the large stretches of noncoding DNA characteristic of eukaryotic chromosomes (Chapter 13). The streamlining of the bacterial genome confers certain benefits. For example, bacteria can reproduce rapidly when nutrients required for growth are available in the local environment. Bacteria replicate their DNA from only one or a small number of initiation sites, so genome size can influence rate of reproduction.
Bacteria do not undergo meiotic cell division and cell fusion. That is, they lack the sexual processes characteristic of eukaryotic organisms (Chapter 11). Despite this, bacterial populations display remarkable genetic diversity. For example, different strains of Pseudomonas aeruginosa, a common disease-causing organism, may differ in genome size by nearly a factor of 2 (3.7 million base pairs compared to 7.1 million base pairs), yet these strains are quite similar in function.
How does this variation arise? In eukaryotic organisms, genes generally pass from parent to offspring. Bacteria also inherit most of their genes from parental cells. However, bacterial cells can also obtain new genes from distant relatives. Called horizontal gene transfer, this process is a major source of genetic diversity in bacteria.
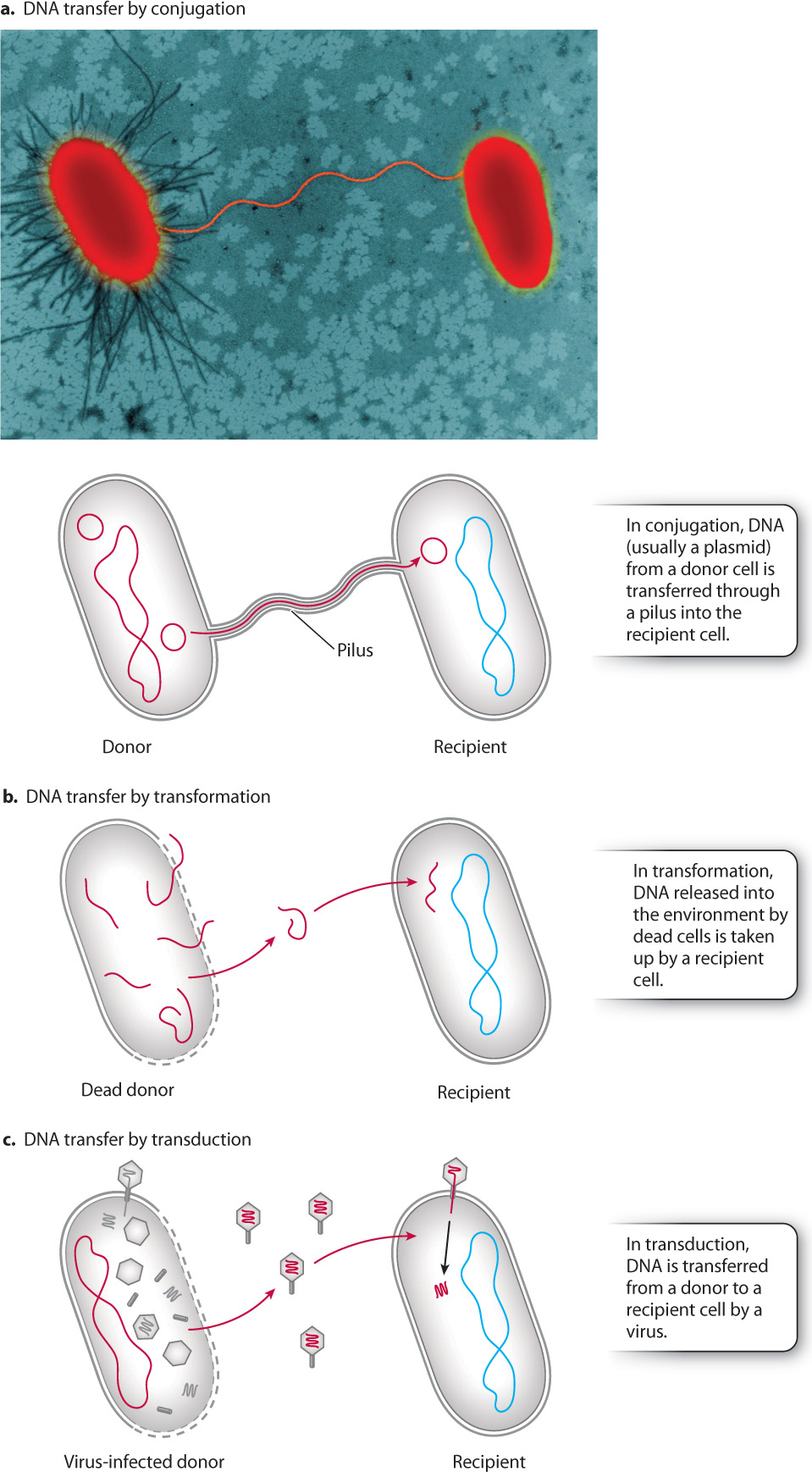
How does DNA move from one bacterial cell into another independently of cell division? Some bacteria synthesize thin strands of membrane-bound cytoplasm called pili that connect them to other cells (Fig. 26.4a). A pilus provides a migration route for the direct cell-to-cell transfer of DNA. This process, called conjugation, commonly transfers plasmids from one cell to another, spreading novel genes throughout a population. Genes that confer resistance to antibiotics are a well-studied example of horizontal gene transfer by conjugation. Biologists interested in genetic engineering use conjugation in the laboratory to introduce genes into cells. But conjugation is just the beginning of genetic exchange in bacteria.
Genes can be transferred from one cell to another without any direct bridge between the cells. DNA released to the environment by cell breakdown can be taken up by other cells, a process called transformation (Fig. 26.4b). Transformation was revealed when experiments showed that harmless strains of the bacteria causing pneumonia could be transformed into virulent strains by exposure to media containing dead cells of disease-causing strains (Chapter 3). Scientists reasoned that the transformation occurred because some substance was being taken up by the living bacteria. The “transforming substance” was later shown to be DNA.
26-5
Viruses provide a third mechanism of horizontal gene transfer. Recall from Chapter 19 that viruses in bacterial cells sometimes integrate their DNA into the host bacterial DNA and persist within the cells as they grow and divide. Before leaving the cell to infect others, the viral DNA removes itself from the bacterial genome. This excision is not always precise, and sometimes additional genetic material from the bacterial host is incorporated into the virus. Viruses released from their host cell go on to infect others, bringing host-derived genes with them. Horizontal gene transfer by means of viruses is called transduction (Fig. 26.4c). It is common in nature and is also widely used in the laboratory to to introduce novel genes into bacteria for medical research.
Horizontal gene transfer allows bacterial cells to gain beneficial genes from organisms distributed throughout the bacterial domain and beyond. Indeed, bacteria everywhere are constantly reshaping their genomes. Bacteria, therefore, are not the poor cousins of eukaryotes, unable to generate genetic diversity by sexual recombination. Instead, bacteria have highly efficient mechanisms for adding and subtracting genes that permit them to evolve and adapt rapidly to local conditions. Perhaps the most widely discussed and worrisome manifestation of horizontal gene transfer is the rapid spread of antibiotic resistance among bacteria (Chapter 48).
Question Quick Check 1
yMwY2YWaAEygcpWpwWU/filbSbqhmdomyTYdEsz7O6vIHp9D49gSxehUk3zkvLxC+x6k3mRHM8fQAiCU5tA0jQJpLyymXUQHmfbkWHzzDy8yeENo0lh13MDpsgpJu4GYCrv42G30FQvNYX4q4wPaoQ/vcgjcp74oBIXYcGK46qrEXIAE9Trsa+G4adxUFF3iWJZD1j2BDeonEDtq/bDTX4Mak0Mnrr1Z8Yzkw/kmArk95YlL4k1RBPlqrJQtbUce26.1.4 The Archaea form a second prokaryotic domain.
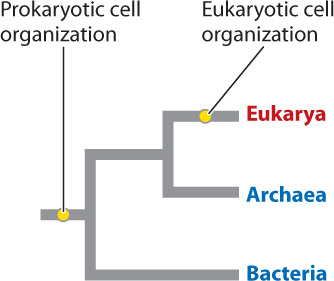
For many years, all prokaryotic cells were classified as bacteria. However, in 1977, George Fox and Carl Woese published a revolutionary hypothesis. From comparisons of RNA molecules from the small subunits of ribosomes (and later, comparisons of the genes for these RNAs), Fox and Woese argued that prokaryotic organisms actually fall into two distinct groups, as different from each other as either is from eukaryotes. If this is true, the tree of life must have three great branches: the Bacteria, the Eukarya, and the limb recognized by Fox and Woese, called the Archaea (Fig. 26.5). Research since the late 1970s confirms this hypothesis. Like bacterial cells, archaeal cells are prokaryotic—they have no membrane-bound nucleus and their genes are arrayed along a single circular chromosome. Moreover, cell size in Archaea is limited by diffusion, and genetic diversity is promoted by horizontal gene transfer, much as in Bacteria.
In detail, however, the Archaea are quite distinctive (Table 26.1). Their membranes are made from lipids different from the fatty acids found in bacterial and eukaryotic membranes. Archaea also show a diversity of molecules in their cell walls, but none has the peptidoglycan characteristic of Bacteria or the cellulose and chitin found in most eukaryotic cell walls.
| Table 26.1: Principal Differences among Archaea, Bacteria, and Eukarya | |||
|---|---|---|---|
| CHARACTERISTIC | ARCHAEA | BACTERIA | EUKARYA |
| Cell contains a nucleus and other membrane-bound organelles | No | No | Yes |
| DNA occurs in a circular form* | Yes | Yes | No |
| Ribosome size | 70S | 70S | 80S |
| Membrane lipids ester-linked† | No | Yes | Yes |
| Photosynthesis with chlorophyll | No | Yes | Yes |
| Capable of growth at temperatures greater than 80°C | Yes | Yes | No |
| Histone proteins present in cell | Yes | No | Yes |
| Operons present in DNA | Yes | Yes | No |
| Introns present in most genes | No | No | Yes |
| Capable of methanogenesis | Yes | No | No |
| Sensitive to the antibiotics chloramphenicol, kanamycin, and streptomycin | No | Yes | No |
| Capable of nitrogen fixation | Yes | Yes | No |
| Capable of chemoautotrophy | Yes | Yes | No |
26-6
Archaea differ from Bacteria in another intriguing way. DNA transcription in archaeons employs RNA polymerase and ribosomes more similar to those of eukaryotes than to bacteria (Chapter 3). Moreover, many of the antibiotics that target protein synthesis in bacteria are ineffective against archaeons, suggesting fundamental differences in translation as well.
Many of the microorganisms first identified as Archaea have unusual physiological properties or inhabit extreme environments. For example, some archaeons live in acid mine water at pH 1 (or less!). Others live in water salty enough to precipitate NaCl (Fig. 26.6), or in deep-sea hydrothermal vents where temperatures can exceed 100ºC. We now know, however, that many archaeons live under less extreme conditions in soils, lakes, and the sea. Indeed, they may be the most abundant organisms throughout much of the ocean.
