27.1 THE EUKARYOTIC CELL: A REVIEW
Chapter 5 introduced the fundamental features of eukaryotic cells. Here, we revisit those attributes (Fig. 27.1) and consider their consequences for function and diversity. How do eukaryotic cells differ from bacteria and archaeons, and how do these differences explain the roles that eukaryotes play in modern ecosystems?
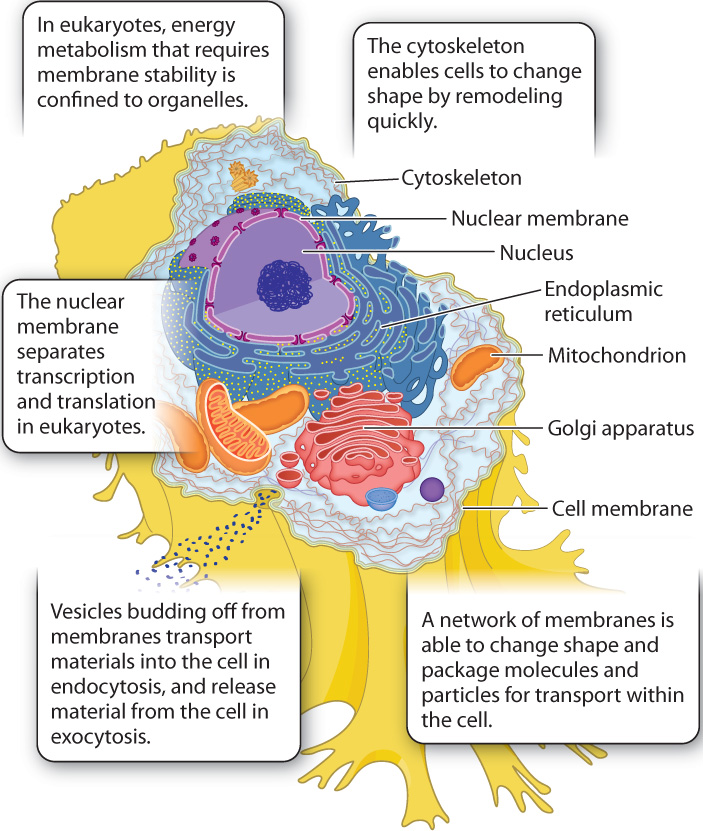
27.1.1 Internal protein scaffolding and dynamic membranes organize the eukaryotic cell.
All cells require a mechanism to maintain spatial order in the cytoplasm. As we saw in Chapter 26, bacteria and archaeons rely primarily on walls that support the cell from the outside, along with a framework of proteins within the cytoplasm. Eukaryotes also use an internal scaffolding of proteins, mostly filaments of actin and microtubules, to organize the cell. This cytoskeleton (Chapter 10), found in all eukaryotic cells, differs in one key property from the rigid protein framework of bacteria: It can be remodeled quickly, enabling cells to change shape.
This characteristic provides some eukaryotes with new possibilities for movement, and it permits them to engulf particles, including other cells, something that prokaryotic cells simply cannot do. Packaged in membrane-lined vesicles, the particles can be transported and processed inside the cell by endocytosis. The same process also works in reverse, and it is then called exocytosis. In exocytosis, vacuoles package newly formed molecules or cytoplasmic waste in vesicles and move them to the cell surface for excretion (Chapter 5). These vesicles and the molecules they carry are transported through the cytoplasm by means of molecular motors associated with the cytoskeleton. In this way, molecules move through the cell at speeds much greater than diffusion would allow. A major consequence is that eukaryotic cells can be much larger than most bacteria.
27-2
Dynamic cytoskeletons require dynamic membranes, and eukaryotes maintain within their cells a remarkable network of membranes called the endomembrane system (Chapter 5). This network includes the nuclear envelope, an assembly of membranes that runs through the cytoplasm called the endoplasmic reticulum (ER) and Golgi apparatus, and a cell or plasma membrane that surrounds the cytoplasm. Moreover, small pieces of membrane can bud off to form tiny vesicles that package molecular signals, nutrients, and wastes within the cell.
All membranes of the endomembrane system are interconnected, either directly or by the movement of vesicles. Many are also capable of changing shape rapidly. In fact, the different membranes are interchangeable in the sense that material originally added to the endoplasmic reticulum may in time be transferred to the cell membrane or nuclear membrane. Biologists like to say that the membranes of eukaryotic cells are in dynamic continuity. Membrane stability, required for energy metabolism, is confined to the mitochondria and chloroplasts.
27.1.2 In eukaryotic cells, energy metabolism is localized in mitochondria and chloroplasts.
We have already noted that, relative to prokaryotic organisms, eukaryotes are fairly limited in the ways they obtain carbon and energy. Moreover, the metabolic processes that power eukaryotic cells take place only in specific organelles—aerobic respiration in the mitochondrion (Chapter 7) and photosynthesis in the chloroplast (Chapter 8). Only limited anaerobic processing of food molecules takes place within the cytoplasm.
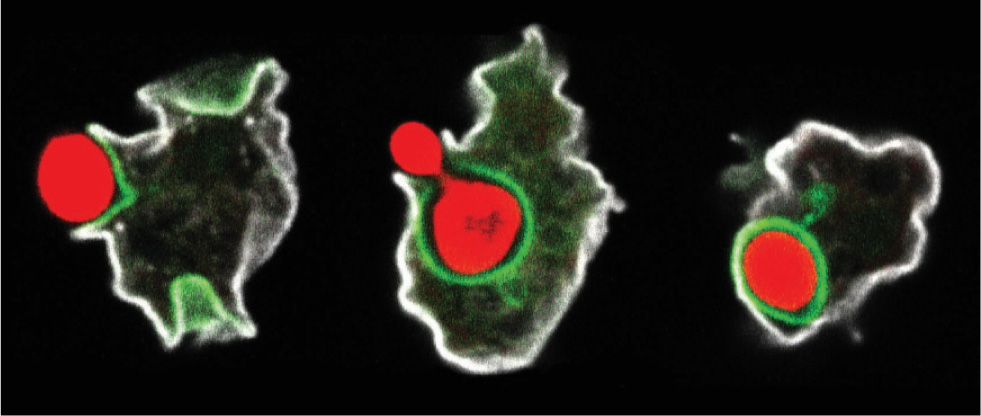
Because energy metabolism is confined to these organelles, eukaryotes free the rest of the cell for other, complementary functions. Unlike bacteria, which absorb individual molecules for respiration, single-celled eukaryotic heterotrophs can engulf particulate food, including other cells (Fig. 27.2). The cells engulf food particles and package them inside a membrane vesicle, which is then transported into the cytoplasm in a process called phagocytosis. Within the cytoplasm, enzymes break down the particles into molecules that can be processed by the mitochondria. Animals can ingest larger foodstuffs, including other animals and plants. In consequence, eukaryotes can exploit sources of food not readily available to bacterial heterotrophs, which feed on individual molecules. This ability opens up a great new ecological possibility—predation—increasing the complexity of interactions among organisms.
The functional flexibility of eukaryotic cells also allows photosynthetic eukaryotes to interact with their environment in ways that photosynthetic bacteria cannot. Unicellular algae (which are eukaryotes) can move effectively through the water column vertically as well as horizontally and therefore can seek and exploit local patches of nutrients. Diatoms, discussed shortly, go one step further. Large internal vacuoles allow them to store nutrients when they are plentiful for later use. Plants, of course, can capture sunlight many meters above the ground, and so have a tremendous advantage on land, as long as their leaves can obtain water and nutrients from the soil in which the plants are rooted.
Taken together, the dynamic cytoskeleton and membrane system, and the related compartmentalization of basic energy metabolism, go a long way toward explaining why eukaryotic cells have so many shapes and prokaryotes have so few.
27.1.3 The organization of the eukaryotic genome also helps explain eukaryotic diversity.
The prokaryotic genome has evolved in such a way that it can replicate quickly. Bacteria and Archaea absorb available nutrients quickly, so both rapid deployment of metabolic proteins and reproduction are key to ecological and evolutionary success. As the majority of a prokaryote’s DNA is arrayed in a single circular chromosome, speed of replication allows for speed of reproduction. As a result, selection favors those strains of Bacteria and Archaea that retain only the genetic material vital to the organism.
27-3
Eukaryotes have multiple linear chromosomes and can begin replication from many sites on each one, enabling them to replicate multiple strands of DNA simultaneously and rapidly (Chapter 12). This ability relieves the evolutionary pressure for streamlining, allowing eukaryotic genomes to build up large amounts of DNA that do not code for proteins. Most of this additional DNA was originally considered to have no function and, indeed, was called “junk DNA.” That view, however, has lost ground in recent years, as the complete nature of the genome has come to be better understood (Chapter 13). At least some of the DNA that does not code for proteins has functions in gene regulation (Chapter 19). This regulatory DNA gives eukaryotes the fine control of gene expression required for both multicellular development and complex life cycles, two major features of eukaryotic diversity.
The evolutionary success of eukaryotes really rests on a combination of features. The innovations of dynamic cytoskeletal and membrane systems gave eukaryotes the structure required for larger cells with complex shapes and the functional ability to ingest other cells. Thus, early unicellular eukaryotes did not gain a foothold in microbial ecosystems by outcompeting bacteria and archaeons. Instead, they succeeded by evolving novel functions. Along with the capacity to remodel cell shape, eukaryotes evolved complex patterns of gene regulation, which in turn enabled unicellular eukaryotes to evolve complex life cycles and multicellular eukaryotes to generate multiple, interacting cell types during growth and development. These abilities opened up still more possibilities for novel functions, which we explore in this and later chapters.
Question Quick Check 1
RGoLrbNjVEBazNIvSE9p5xBA/tHgytcJYPn43TEVgXbrvxheRVLNHJ7xU8V+Xhk/euy+tGcl6Yh5Fk4d1NYzNiWyOUHLpXJKGRch4C0EiCLl8FClOs1unzW9dMNTLNa/aLRyfrLDl6Z0y04FGf0anhjaLfpTMW2XzjFlwVEMOAc=27.1.4 Sex promotes genetic diversity in eukaryotes and gives rise to distinctive life cycles.
In Chapter 26, we saw how Bacteria and Archaea generate genetic diversity by means of horizontal gene transfer. Horizontal gene transfer has been documented in eukaryotic species, but it is relatively uncommon. How then do eukaryotes build and maintain genetic diversity within populations? The answer is sex.
Sexual reproduction involves the formation of gametes by meiosis and their fusion by fertilization (Chapters 11 and 42). Sex promotes genetic variation in two simple ways. First, meiotic cell division results in daughter cells (gametes or spores) that are genetically unique—that is, the combinations of alleles in each gamete are different from each other and from the parental cell—as a result of recombination and independent assortment. Second, in fertilization, new combinations of genes are brought together by the fusion of gametes. Interestingly, a few eukaryotic groups have lost the capacity for sexual reproduction. In the best-studied of these eukaryotes, tiny animals called bdelloid rotifers, genetic diversity is actually high, maintained by high rates of horizontal gene transfer. Apparently, horizontal gene transfer and sexual reproduction are alternative and, to a large extent, mutually exclusive ways of maintaining genetic diversity within populations.
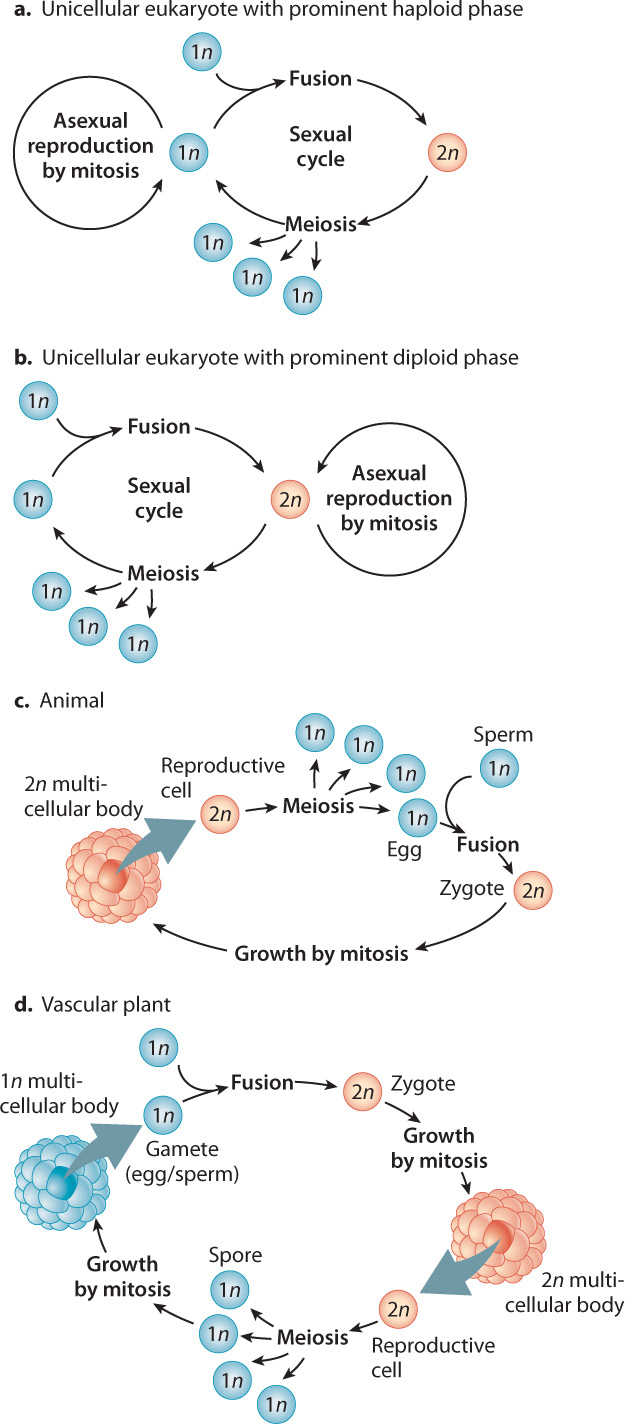
As we saw in Chapter 11, meiotic cell division results in cells with one set of chromosomes. Such cells are haploid. Sexual fusion brings two haploid (1n) cells together to produce a diploid (2n) cell that has two sets of chromosomes. The life cycle of sexually reproducing eukaryotes, then, necessarily alternates between haploid and diploid states.
Many single-celled eukaryotes normally exist in the haploid stage and reproduce asexually by mitotic cell division (Fig. 27.3a). An example is provided by the green alga Chlamydomonas. Typically, starvation or other environmental stress can cause two cells to fuse, forming a diploid cell, or zygote. The zygote formed by these single-celled eukaryotes commonly functions as a resting cell. It covers itself with a protective wall and then lies dormant until environmental conditions improve. In time, further signals from the environment induce meiotic cell division, resulting in four genetically distinct haploid cells that emerge from their protective coating to complete the life cycle.
As shown in Fig. 27.3b, some single-celled eukaryotes normally exist as diploid cells. For example, in diatoms, single-celled eukaryotes commonly found in lakes, soils, and the oceans, cells are mostly diploid and reproduce asexually by mitotic cell division to make more diploid cells. Because of the constraints of their mineralized skeletons, diatoms become smaller with each asexual division. Once a critical size is reached, meiotic cell division is triggered, producing haploid gametes that fuse to regenerate the diploid state as a round, thick-walled cell. This cell grows and then germinates to form actively growing, skeletonized cells. In diatoms, then, short-lived gametes constitute the only haploid phase of the life cycle.
27-4
The two life cycles just introduced are similar in many ways. Both involve haploid cells that fuse to form diploid cells, and diploid cells that undergo meiotic cell division to generate haploid cells. Both life cycles also commonly include cells capable of persisting in a protected form when the environment becomes stressful. Why some single-celled eukaryotes usually occur as haploid cells and others generally occur as diploid cells remains unknown—a good question for continuing research.
Sexual reproduction has never been observed in some eukaryotes, but most species appear to be capable of sex, even if they reproduce asexually most of the time. Variations on the eukaryotic life cycle can be complex, especially in parasitic microorganisms that have multiple animal hosts. All, however, have the same fundamental components as the two life cycles described here.
In succeeding chapters, we discuss multicellular organisms in detail. Here we note only that animals, plants, and other complex multicellular organisms have life cycles with the same features as those just discussed. The big difference is that in animals, the zygote divides many times to form a multicellular diploid body before a small subset of cells within the body undergoes meiotic cell division to form haploid gametes (eggs and sperm). During fertilization, the egg and sperm combine sexually to form a zygote (Fig. 27.3c). In animals, as in diatoms, the only haploid phase of the life cycle is the gamete. As we will see in Chapter 30, plants have two multicellular phases in their life cycle, one haploid and one diploid (Fig. 27.3d). Like single-celled eukaryotes, many plants and animals can reproduce asexually. Humans and other mammals are unusual in that we have no capacity for asexual reproduction.