28.1 THE PHYLOGENETIC DISTRIBUTION OF MULTICELLULAR ORGANISMS
As we discussed in Chapter 26, most prokaryotic organisms are composed of a single cell, although some form simple filaments or live in colonies. A few types of bacterium, notably some cyanobacteria, develop several different cell types. No bacteria, however, develop macroscopic bodies with distinct types of tissues. Only eukaryotes have evolved those.
28.1.1 Simple multicellularity is widespread among eukaryotes.
A recent survey of eukaryotic organisms recognized 119 major groups within the superkingdoms discussed in Chapter 27. Of these, 83 contain only single-celled organisms, predominantly cells that engulf other microorganisms or ingest small organic particles, photosynthetic cells that live suspended in the water column, or parasitic cells that live within other organisms. Each of the 36 remaining branches exhibits some cases of simple multicellularity, mostly in the form of filaments, hollow balls, or sheets of little-differentiated cells (Fig. 28.1).
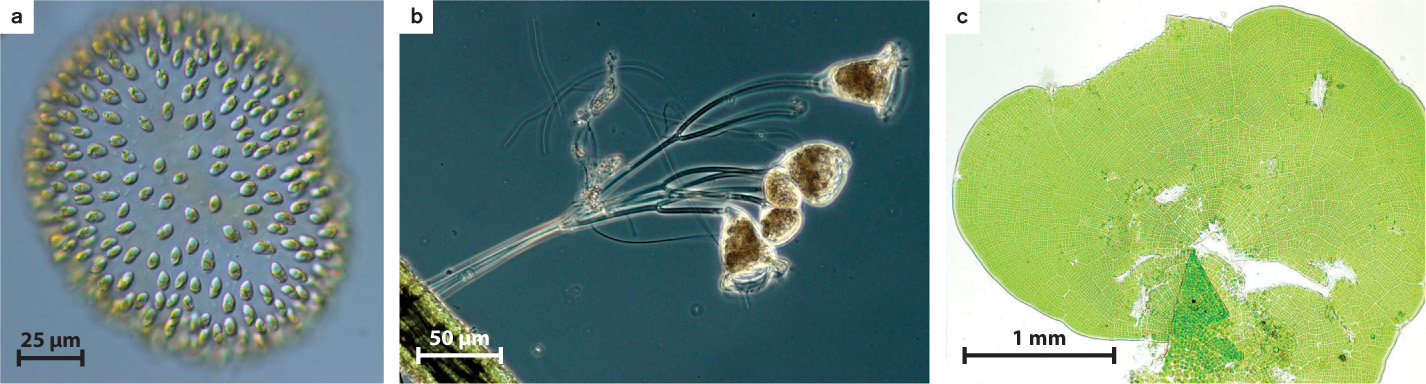
Simple multicellular eukaryotes share several properties. In many, adhesive molecules cause adjacent cells to stick together, but there is relatively little communication or transfer of resources between cells and little differentiation of specialized cell types. Most or all of the cells in simple multicellular organisms retain a full range of functions, including reproduction, and so the penalty paid by the organism for individual cell death is usually small. Importantly, in simple multicellular organisms, every cell is in direct contact with the external environment, at least during phases of the life cycle when the cells must acquire nutrients.
28-2
Most simple multicellular species are forms of algae, although stalklike colonies of particle feeders have evolved at least three times, and simple filamentous fungi absorb organic molecules as sources of carbon and energy. In addition, four eukaryotic groups have achieved simple multicellularity by a different route, aggregating during just one stage of the life cycle. Slime molds are the best-known example (Chapter 27).

Six other groups (two algal and three protozoan, plus a group of fungi) have evolved what is called coenocytic organization. In these organisms, the nucleus divides multiple times, but the nuclei are not partitioned into individual cells. The result is large cells—sometimes even visible to the naked eye—with many nuclei. The green algae Codium and Caulerpa are common examples of coenocytic organisms found along the shorelines of temperate and tropical seas, respectively (Fig. 28.2). Coenocytic rhizarians on the deep seafloor can be 10 to 20 cm long. There is no evidence that any of these groups evolved from truly multicellular ancestors, nor have any given rise to complex multicellular descendants.
Why did simple multicellular organisms evolve? One selective advantage is that multicellularity helps organisms to avoid protozoan predators. In an illuminating experiment, single-celled green algae were grown in the presence of a protistan predator. Within 10 to 20 generations, most of the algae were living in eight-cell colonies that were essentially invulnerable to predator attack. Another advantage is that multicellular organisms may be able to maintain their position on a surface or in the water column better than their single-celled relatives. Seaweeds, for example, live anchored to the seafloor in places where light and nutrients support growth. And in colonial heterotrophs such as the stalked ciliate Epistylis (see Fig. 28.1b), the coordinated beating of flagella assists feeding by directing currents of food-laden water toward the cells. As we will see, complex multicellular organisms evolved from simple multicellular ancestors, but most taxonomic groups containing simple multicellular organisms never gave rise to complex descendants.
28-3
28.1.2 Complex multicellularity evolved several times.

Complex multicellular organisms mark the biological world of our daily existence. Plants, animals, and, if you look a little more closely, mushrooms and complex seaweeds define our perceptions of landscapes and the coastal ocean (Fig. 28.3). Plants and animals differ in many ways, but they have some fundamental features in common. Both have highly developed molecular mechanisms for adhesion between cells, and they have specialized structures that allow cells to communicate with one another. Plants and animals also both display complex patterns of cellular and tissue differentiation, guided by networks of regulatory genes. All complex multicellular organisms exhibit these properties. In general, in complex multicellular organisms only a small subset of all cells contributes to reproduction. Cell or tissue loss can be lethal for the entire organism. Notably, complex multicellular organisms have a three-dimensional organization, so only some cells are in direct contact with the environment.
The presence of both exterior and interior cells is an important characteristic of complex multicellular organisms. Because some cells are buried within tissues, relatively far from the exterior of the organism, they do not have direct access to nutrients. Therefore, interior cells cannot grow as fast as surface cells unless there is a mechanism to transfer resources from one cell to another. Also, interior cells do not receive signals directly from the environment, even though all cells must be able to respond to environmental signals if the organism is to grow, reproduce, and survive. Complex multicellular organisms, therefore, require mechanisms for transferring environmental signals received by cells at the body’s surface to interior cells, where genes will be activated or repressed in response. That process has a familiar ring to it. Development in complex multicellular organisms can be defined as increasing or decreasing gene expression in response to molecular signals from surrounding cells (Chapter 20).
28-4
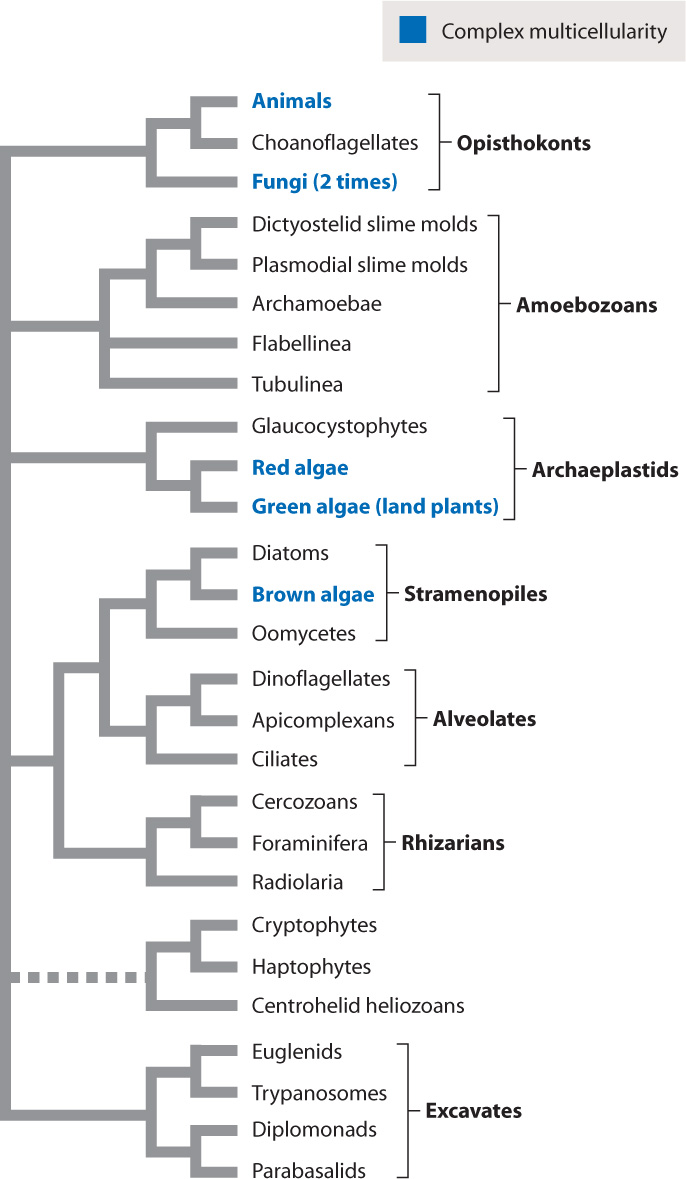
Complex multicellularity evolved at least six separate times in different eukaryotic groups (Fig. 28.4). Obviously, complex multicellularity characterizes animals, but it also evolved at least twice in the fungi, once in the green algal lineage that gave rise to land plants, once in the red algae, and once in the brown algae, producing the giant kelps that form forests in the sea.