31.1 SHOOT GROWTH AND DEVELOPMENT
In tropical forests, vines weave their way upward through the branches of trees. Many of these vines have long, thin stems with widely spaced leaves (Fig 31.1a). In contrast, a barrel cactus, living in the desert, has thick stems and leaves modified to form sharp spines (Fig. 31.1b). Although they look nothing alike, these two plants are constructed in the same way. Plant shoots are modular, meaning that they are formed of repeating units. Each unit consists of a node, the point where one or more leaves are attached, and an internode, the segment between two nodes (Fig. 31.1c). In vines the internodes are long and the leaves large, whereas in cacti the internodes are short and the leaves thin and sharp. The modular nature of plants helps us understand how the capacity for continued growth gives rise to the tremendous variation in plant form that we see around us.

31.1.1 Shoots grow by adding new cells at their tips.
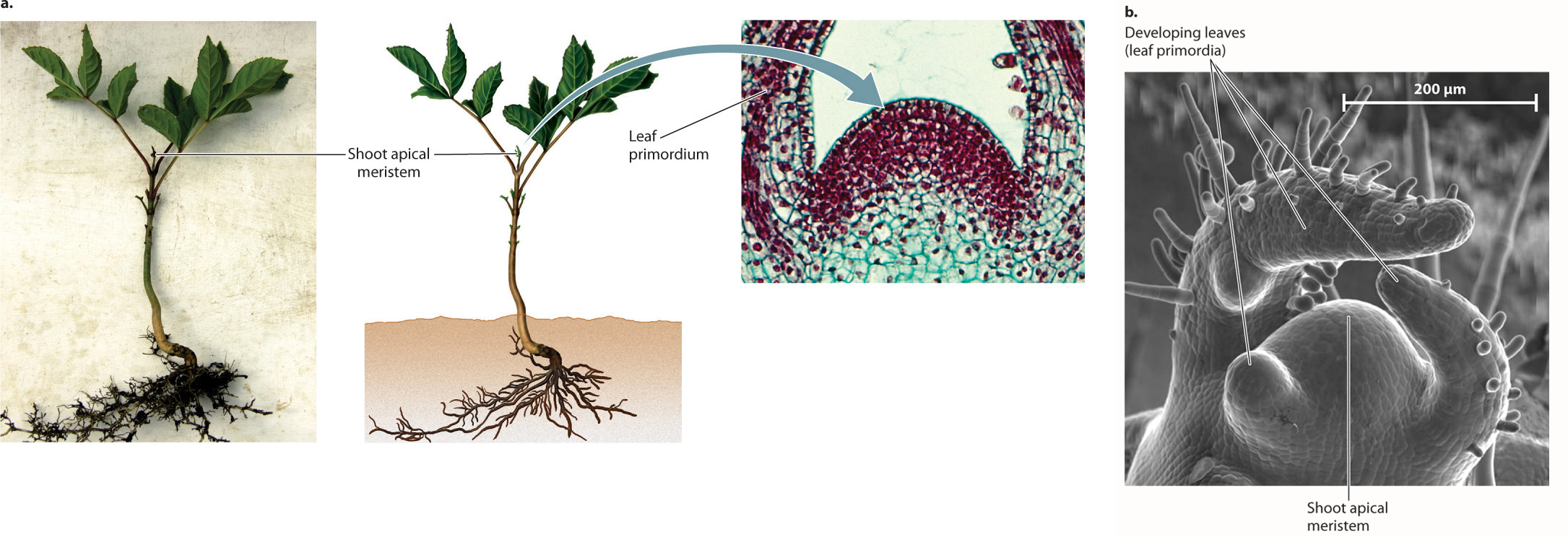
At the very tip of each branch, commonly hidden beneath an array of young leaves called leaf primordia (singular, primordium), lies a tiny dome of cells called the shoot apical meristem (Fig. 31.2). The shoot apical meristem is a group of totipotent cells that, like embryonic stem cells in animals, gives rise to new tissues. As we will see, plants have meristems in several parts of their bodies, but upward growth occurs exclusively in and just below the shoot apical meristem. This is where cell division occurs, generating all the new cells that serve to elongate stems. The shoot apical meristem also initiates leaves and produces new meristems, which allow plants to branch.
31-3
An important characteristic of the shoot meristem is that it maintains a constant size even though it is a region of active cell division. As new cells are added near the shoot tip, cells that are farther from the shoot tip cease to divide. This means that the cells of the meristem are constantly being lost and replaced. Cells near the shoot tip maintain their totipotency by the expression of meristem identity genes, which contribute to meristem stability and function. The expression of meristem identity genes is controlled by a network of chemical signals produced by cells at the very tip of the stem. Because these signals can be transmitted only over a limited distance, only the cells close to the shoot tip express meristem identity genes.
Question Quick Check 1
XiSj9Nv9wB9qCiw3qFmfAp7liyx9T0ZOw9L/N4DAd9CzqfpNcHdGN/hR7BPwDeoZ9yeKxMfnHwn2pybG31.1.2 Stem elongation occurs primarily in a zone just below the apical meristem where new cells elongate.
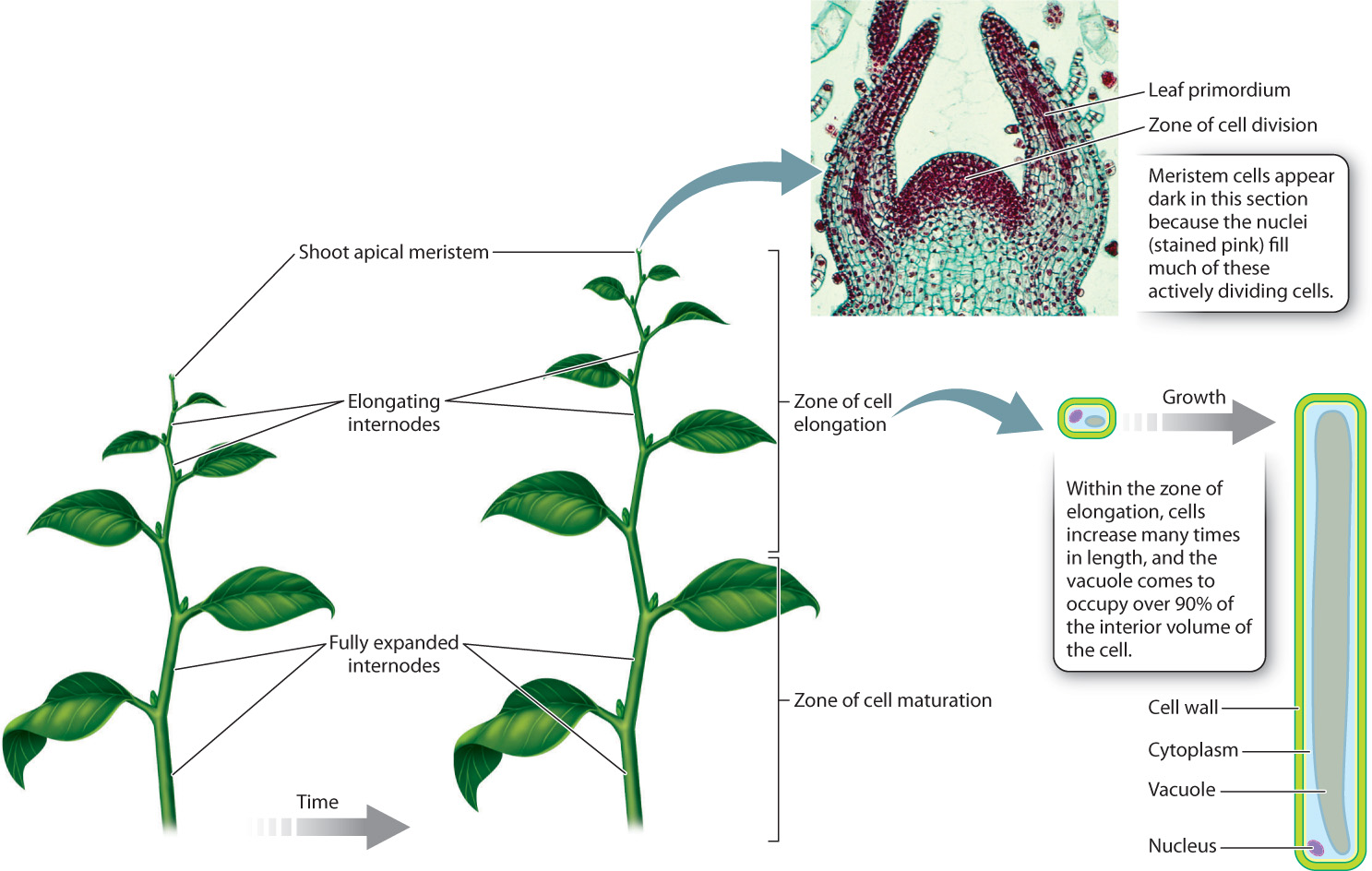
In animals, cell division and cell enlargement typically go hand in hand. In plants, most of the increase in cell size occurs after mitotic cell division is complete. Cells that are too far from the shoot tip to express meristem identity genes cease to divide, yet they continue to grow. This results in a zone of cell elongation located just beneath the shoot apical meristem (Fig. 31.3). It is here that most of the elongation of stems occurs. If you imagine all the cells in the stem of a 100-m redwood tree at the size they were when they left the zone of cell division—the tree would be less than 5 m tall.
31-4
In the zone of cell elongation, each cell grows many times more in length than in width. The reason is that the strong cellulose microfibrils wrapped around the cell make it difficult for the cell to expand in girth. The large central vacuole that characterizes mature plant cells forms when the cell is in the zone of cell elongation. In fact, most of the increase in cell volume is due to the uptake of water and solutes that fill the vacuole. This explains in part why plant growth is markedly reduced during periods of drought.
As we move even farther from the shoot tip, cells reach their final size and complete their differentiation into the mature cell types in leaves and stems. This organization into successive zones of cell division, cell elongation, and cell maturation allows stems to grow without any predetermined limit to their length. It also means that we can follow the time course of development by moving along the stem from the tip toward the base.
Once cells have matured, they no longer expand. Thus, during the upward growth of the shoot, it is the production and elongation of cells at the shoot tip that lift the meristem ever higher into the air. Imagine that, as a 10-year-old, you carved your initials into a tree. Twenty years later, your initials will be exactly the same distance from the ground as they were on the day you inscribed them. The letters, however, will be wider—a phenomenon we discuss later in this chapter.
31.1.3 The shoot apical meristem controls the production and arrangement of leaves.
In most plants, leaves are the principal sites of photosynthesis. Because light is needed for photosynthesis, the arrangement of the leaves along a stem has a major impact on their function. Each species has a characteristic number of leaves attached at each node along the stem. Some species have only a single leaf at each node, whereas others have two or more. How the leaves are positioned around the stem varies in a predictable fashion (Fig. 31.4). The regular placement of leaves reduces the shading of one leaf by another and thus enhances the ability of plants to obtain sunlight.


Leaves begin as small bumps, the leaf primordia, which form on the sides of the shoot apical meristem (see Fig. 31.2). The regular arrangement of leaves around the stem is controlled by the fact that each successive leaf primordium is located as far away as possible from all previously formed primordia. One hypothesis proposed to explain this developmental pattern is that the diffusion of chemical signals from developing leaf primordia creates regions where the growth of new primordia is inhibited. The result is an arrangement that prevents leaves from being produced one on top of another.
As noted previously, the earliest vascular plants were simple branching stems, and photosynthesis took place along the length of the shoot. As evolution proceeded, however, some branch systems became flattened, their axes growing in a plane that facilitated the capture of light. These planar branches lost the capacity for continued growth. By about 380 million years ago, the planar branches became modified into structures recognizable as leaves (Fig. 31.5).
The evolution of leaves required three developmental changes. First, the genetic program to produce a three-dimensional stem became modified to form flattened organs. Second, apical meristem identity genes were down-regulated and leaf identity genes up-regulated, resulting in a specialized organ incapable of continuing growth. And third, new meristems evolved, enabling leaves to expand into flattened photosynthetic structures that capture sunlight. In fern leaves, these meristems are located along the leaf margin. In pine needles, they occur at the base of each needle. In flowering plants, meristematic cells can be distributed throughout the developing leaf, making possible a diversity of leaf shape. In contrast to cells in the shoot apical meristems which can divide continuously throughout the lifetime of a plant, leaf meristematic cells divide only for a relatively short period of time. This explains why leaves grow to a final size.
When we think of leaves, it is the green photosynthetic ones that first come to mind. However, many plants produce leaves that are specialized for functions other than photosynthesis, including climbing, trapping insects, and attracting pollinators (Fig. 31.6). Plants that overwinter produce bud scales that protect shoot apical meristems from desiccation and damage due to cold. Bud scales may not look like leaves, but they form from leaf primordia and are arranged in the same way around the stem as the green leaves produced in spring.

31-5
31.1.4 The development of new apical meristems allows stems to branch.
Vascular plants evolved the ability to branch even before they evolved either roots or leaves. Branching was important to these first plants because it allowed them to produce more sporangia. Branching allows present-day plants to support greater numbers of both reproductive structures and leaves.
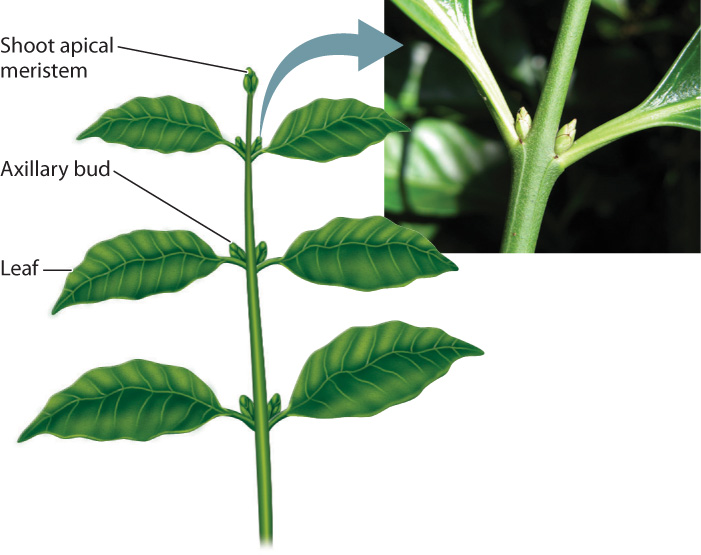
In lycophytes and in ferns and horsetails, branching occurs when the shoot apical meristem divides in two, giving rise to two stems. In seed plants, branches grow out from axillary buds (also called lateral buds), which are meristems that form at the base of each leaf (Fig. 31.7). Axillary buds have the same structure and developmental potential as the apical meristem and express the same meristem identity genes. However, the axillary buds remain dormant until triggered to grow, remaining attached to the stem even after leaves are shed. Thus, axillary buds provide seed plants with many points along their stem where new branches can form.
31-6
31.1.5 Flowers grow from and consume shoot meristems.
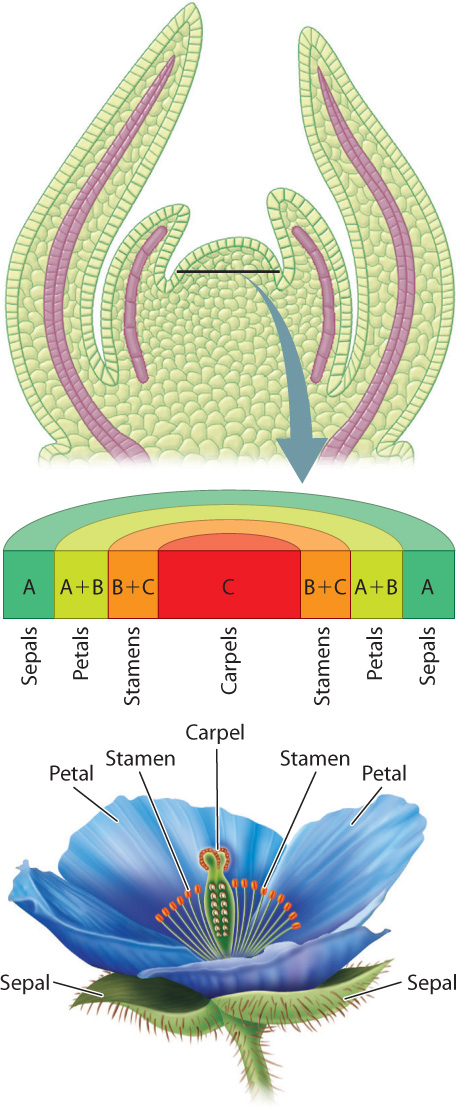
Just as leaves and branches grow from meristems, so, too, do flowers. Flowers can be produced at the tip of a plant or shoot as a result of the conversion of the apical meristem into a floral meristem, or at the base of leaves as the products of axillary buds. As with leaves, floral meristems lose their capacity for continued growth. All the cells differentiate entirely during flower development.
As we saw in Chapter 30, flowering is triggered by florigen, a protein produced in leaves and transported through the phloem to apical meristems and axillary buds. Florigen triggers the transition to floral meristems by initiating the down-regulation of meristem identity genes and the up-regulation of genes that govern floral identity. The arrangement of primordia is altered to produce four whorls that give rise to the sepals, petals, stamens, and carpels. Once the meristem is launched along the trajectory for flower development, the identity of each whorl is controlled by the expression of homeotic genes (Chapter 20). Three classes of genes, referred to as A, B, and C, are expressed in overlapping rings around the meristem and serve as master controllers for the development of specific floral organs (Fig. 31.8).