32.3 ALLOCATING RESOURCES TO DEFENSE
By now, it should be clear that plants invest substantial resources in defense, resources that might otherwise be used for growth and reproduction. Given the considerable cost of plant defenses, their benefits must be correspondingly large. Obviously, if a commonly encountered pathogen or herbivore is lethal, the resources invested in defense provide reproductive advantage. But what if the pathogen or herbivore usually damages only half of a plant’s leaves? Or 10%? Or encounters the plant only infrequently?
In nature, the balance between cost and benefit is achieved in several different ways. When a threat is constant or generalized, plants mount constitutive defenses—defenses that are always active. When a threat is uncommon, however, there is an advantage to mounting a defense only when the threat is encountered. A defense that is mounted only when the plant senses the threat is an inducible defense.
32.3.1 Plants can sense and respond to herbivores.
When a plant is attacked by herbivores, it begins to express a new set of genes. Many of these genes increase the production of chemical defenses, while others stimulate the mechanical reinforcement of cell walls. In addition, herbivore damage triggers the synthesis of jasmonic acid, a signal that is transmitted through the phloem. Exposure to jasmonic acid induces the transcription of defensive genes even in parts of the plant not directly affected by herbivores. Methyl-jasmonic acid is highly volatile and can move through the air to reach portions of the plant that are not actively importing carbohydrates through the phloem.
Question Quick Check 3
tRKv43LjPw2dd7wz0EdnFQJye0tFFI9FS0pUBfCMrqf/cVfItrLrTVhSuQ7yYsiX7HBh5cOuep0VwOcM44IycdioKWPBmY6EtXehgLThfnQ=Another way in which plants can sense and respond to herbivores is illustrated by coyote tobacco (Nicotiana attenuata), a relative of tobacco native to dry open habitats of the American West (Fig. 32.15). For more than 15 years, biologists have intensively studied the mechanisms by which N. attenuata defends itself against herbivores, mechanisms that provide a fascinating window into how plants protect themselves under natural conditions.

32-14
Nicotiana attenuata is attacked by a wide variety of herbivores, both generalists and specialists. Which species cause the most damage, however, varies significantly from year to year. This means that N. attenuata must either produce costly defenses for herbivores that might not show up in any particular year or be able to identify and respond to specific threats.
A major component of N. attenuata’s defense against being eaten is the production of the alkaloid nicotine. This defense is metabolically expensive because it contains nitrogen. Nicotine is an effective defense against many generalist herbivores. However, N. attenuata is also attacked by a specialist herbivore, the tobacco hornworm caterpillar, Manduca sexta. These caterpillars, which can reach 7 cm in length, have evolved the ability to sequester and secrete nicotine. By this means, these caterpillars are able to consume leaves with high levels of nicotine. If nicotine is not an effective defense against this voracious herbivore, what can N. attenuata do to protect itself?
It turns out that N. attenuata is able to recognize specific chemicals in the saliva of M. sexta. When damaged by most herbivores—or a pair of scissors—N. attenuata responds by producing nicotine. But when attacked by M. sexta, N. attenuata does not waste time and resources synthesizing nicotine. Instead, it protects itself in an entirely different way, by attracting other insects that will attack its herbivores.
32.3.2 Plants produce volatile signals that attract insects that prey upon herbivores.
How does N. attenuata attract its insect allies? A hint comes from the smell of a new-mown lawn. This distinctive smell originates from chemicals released from the cut blades of grass. Some of the chemicals released when plants are damaged represent nothing more than the exposure of cell interiors to the air. Others, however, are produced specifically in response to damage.
Studies first conducted in the 1980s showed that undamaged plants increase their synthesis of chemical defenses when neighboring plants are attacked by herbivores (Fig. 32.16). Although these plants were described in the popular press as “talking trees,” the plants under attack are not altruistically warning their neighbors. Instead, subsequent studies suggest that these plants are sending signals directed at other insects. When N. attenuata is attacked by M. sexta caterpillars, it produces volatile signals that attract insects that prey on the eggs and larvae of M. sexta. The adage “The enemy of my enemy is my friend” seems to hold true when it comes to protecting plants from herbivore damage. Neighboring plants sense these chemical signals and respond by ramping up their own defenses.
FIG. 32.16: Can plants communicate?
BACKGROUND Plants respond to herbivore damage by producing chemicals that make their tissues less palatable. Field experiments in which caterpillars were added to target plants suggested that undamaged neighboring plants increased their production of defensive chemicals, whereas plants farther away did not.
HYPOTHESIS Volatile chemicals released from plants that have been attacked by herbivores elicit a defensive response in undamaged plants.
EXPERIMENT One set of plants was placed downwind of other plants in which two leaves were torn, simulating herbivore damage, and a set of control plants was placed downwind of an empty chamber. The concentrations of defensive compounds in all sets of plants were measured and compared.
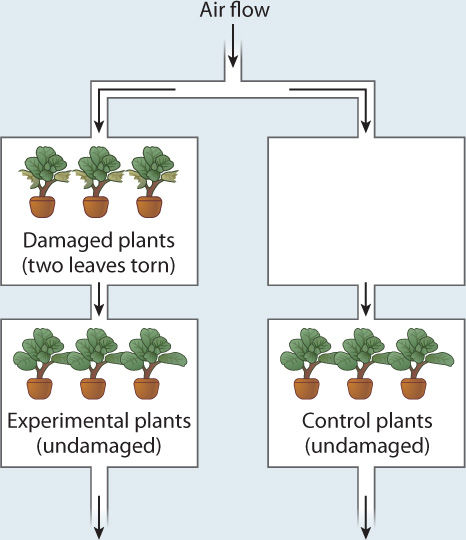
RESULTS When measured 52 hours after the initial damage, both the damaged plants and the undamaged experimental plants exhibited elevated levels of defensive compounds in their leaves compared to control plants.
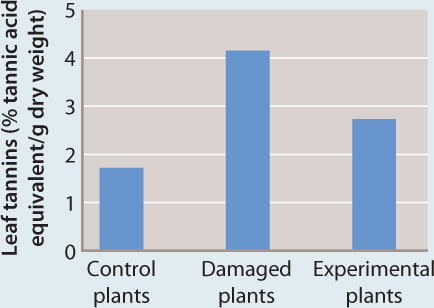
CONCLUSION Volatile chemicals released from damaged plant tissues can trigger the production of defensive chemicals in undamaged plants.
FOLLOW-UP WORK The volatile chemicals released from damaged plants have been identified and shown to attract predatory animals that feed on the herbivores or parasitize their eggs or both. Thus, when plants respond to damaged neighbors they are “eavesdropping” on signals that likely evolved to attract predatory insects, rather than to warn neighboring plants.
SOURCE Baldwin, I. T., and J. C. Schulze. 1983. “Rapid Changes in Tree Leaf Chemistry Induced by Damage: Evidence for Communication Between Plants.” Science 221:277–279.
32.3.3 Nutrient-rich environments select for plants that allocate more resources to growth than to defense.
Plant species vary significantly in the extent to which they allocate resources to defense. Because defenses are costly to produce, you might think that plants growing in nutrient-rich habitats should be the best defended. However, consider the counterargument: Plants growing on nutrient-poor soils might invest heavily in defense because they cannot afford to replace tissues lost to herbivores. Such plants would grow relatively slowly because their resources were being used to build defenses rather than to produce new leaves and roots. By contrast, plants from nutrient-rich habitats would favor growth over defense because they could more readily replace lost or damaged tissues.
32-15
This situation is an example of a trade-off: Something is gained and, at the same time, something is lost. You can’t have your cake and eat it, too—you have to make a choice. With plants, there can be a trade-off between growth and defense in which allocating resources for one means that those resources cannot be allocated for the other.
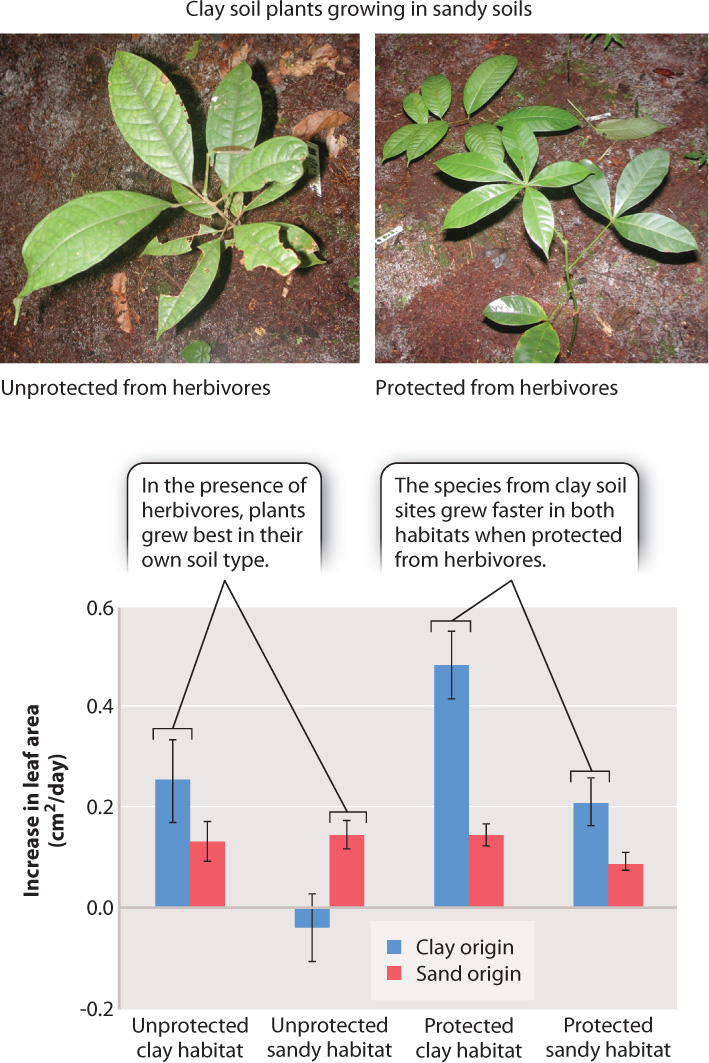
A recent study in the Amazon rain forest has focused on a possible trade-off between growth and defense. Researchers compared species found on nutrient-rich clay soils with closely related species found on nutrient-poor sandy soils. They found that species from the nutrient-rich clay soils invested less heavily in defenses than did species from the sandy sites. Because the clay-soil species divert fewer resources to defense, they grow faster in either soil type compared to sandy-soil species—as long as they are grown under a net that excludes herbivores (Fig. 32.17). However, when grown out in the open, each species grows best in the soil type on which it is naturally found.
The clay-soil species grow poorly on sandy soils when not protected by a net because, without ample soil nutrients available, they are unable to replace tissues consumed by herbivores. Conversely, when grown on the nutrient-rich soils, species from the sandy habitats are poor competitors because they allocate resources that could have been used for growth to defensive compounds and structures. What we see is a classic example of a trade-off.
32-16
Question Quick Check 4
zHcCk34Y4p0etScto+MelQbVoBf/HS7edwdGY+jrGB6vJSG5xSt9UhbsB3B1y3zkt+Hdnq6XtxpE3b4srumpj5cRSV8vdG8t1/ZN27mYC6PBd5r9l3jrDRHZTKwQIxhoR1oMec0UKz4R1oPjwbknl0Zy7f2hplvOSSmKNRkBAZPz/6aX3soKq7vQU9QKpkCzLrvG/Moo/LaSB4O932.3.4 Exposure to multiple threats can lead to trade-offs.
In the real world, plants are routinely confronted with more than one threat. A plant may be attacked simultaneously by both a pathogen and an herbivore, or it might be in danger of being shaded by a fast-growing neighbor at the same time that it must defend its leaves from being eaten. Given that the resources a plant can draw on are limited, how do plants respond to multiple threats?
Often, the response to multiple threats suggests a trade-off. For example, exposure to a pathogen can reduce the ability to respond to a subsequent herbivore attack. Tobacco plants in which systemic acquired resistance has been activated as a result of exposure to tobacco mosaic virus are more susceptible to the hornworm caterpillar, Manduca sexta. Studies indicate that cross talk between the signaling pathways for responding to pathogens versus herbivores is responsible for the reduced ability to respond subsequently to herbivores.
There can also be trade-offs between threats resulting from competition with neighboring plants and attack by herbivores. For example, when the phytochrome receptor in plants detects the presence of neighboring plants (Chapter 31), plants not only allocate more resources to growing tall but also produce fewer defensive chemicals in response to herbivore damage. Once again, interactions among the signaling molecules associated with each of these processes are responsible for mediating the trade-off.