35.4 NERVOUS SYSTEM ORGANIZATION
The brain is considered the body’s command center: It receives sensory information from the eyes, ears, and skin and issues instructions to the rest of the body. Up to this point, we have examined how individual neurons send signals to other nerves and muscles. How are these neurons organized in the body to sense stimuli, process them, and issue an appropriate response? How does the brain coordinate the movement of limbs necessary to walk or run, and adjust heart rate and breathing in response to exercise? In this section, we look at how the nervous system is organized to allow the brain and the body to communicate with each other.
35.4.1 Nervous systems are organized into peripheral and central components.
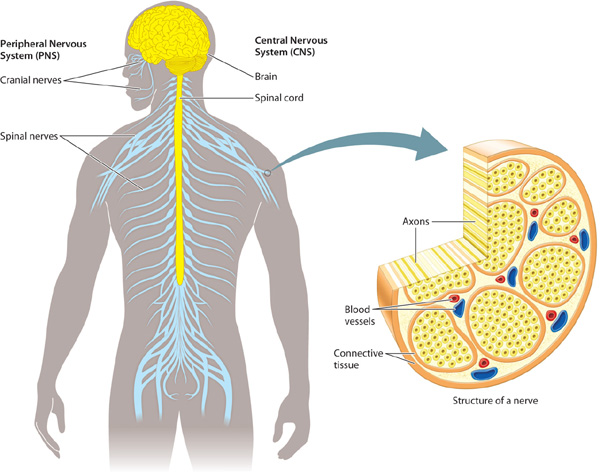
As animals evolved the ability to sense and coordinate responses to increasingly complex stimuli, their nervous systems became organized into peripheral and central components (Fig. 35.17). Not surprisingly, your eyes, sense of touch, and other sensory organs are located on the surface of the body, where they can receive signals from the environment. In contrast, the brain, centrally located ganglia, and a main nerve cord are located in the interior.
35-16
How do structures in the interior and near the surface communicate? Nerves form the lines of communication between these nervous system structures. In general, neuron cell bodies are grouped compactly together in sensory organs, ganglia, and a main nerve cord that extends from the brain. In contrast, nerves are composed mainly of axons from many different nerve cells. For example, the optic nerve contains axons that travel from nerve cells in the eye to the brain.
Sensory and motor nerves make up the peripheral nervous system (PNS). These nerves communicate with the brain, a main nerve cord, or—in the case of organisms such as flatworms that lack a brain—with centralized information-processing ganglia. The brain, main nerve cord, and centralized ganglia together make up the central nervous system (CNS). Information gathered by sensory organs arrives mainly from the animal’s periphery, at or near its body surface. After processing the information, the ganglia and brain send commands back to peripheral nerves that coordinate the activity of muscles and glands in different regions of the animal’s body. This organization ensures that an animal achieves a coordinated behavioral and physiological response to the stimuli received from its environment. Only the diffuse nerve nets of cnidarians lack defined central and peripheral components.
Specialized sensory organs transmit information from the animal’s periphery by afferent neurons, which send information toward the CNS. Efferent neurons send signals away from the CNS, communicating with muscles and other tissues and organs. The peripheral nervous system also includes interneurons and ganglia that integrate and process information in local regions of the animal’s body. For example, vertebrates and invertebrates have ganglia that lie outside each segment of their primary nerve cord. However, the bulk of information processing, particularly in animals that exhibit a greater capacity for learning and memory, occurs in the CNS.
The central nervous system of vertebrates includes both the brain and spinal cord (Fig. 35.17). The spinal cord is a central tract of neurons that passes through the vertebrae to transmit information between the brain and the periphery of the body. The vertebrate spinal cord is divided into segments, each controlling body movement in a particular region along the animal’s length. Each spinal cord segment contains axons from peripheral sensory neurons, a set of interneurons, and a set of motor neuron cell bodies. These are distinct from, but often associated with, segmental ganglia that lie outside the spinal cord.
In humans and other vertebrate animals, the peripheral nervous system is organized into left and right sets of cranial nerves located within the head and spinal nerves running from the spinal cord to the periphery. Most of the cranial nerves and all of the spinal nerves contain axons of both sensory and motor neurons. Cranial nerves link specialized sensory organs (eyes, ears, tongue) to the brain. Cranial nerves also control eye movement, facial expression, speech, and feeding. Some cranial nerves, such as the olfactory and optic nerves, contain only sensory axons. Spinal nerves exit from the spinal cord through openings located between adjacent vertebrae to thread through the trunk and limbs of an animal’s body. These nerves receive sensory information from receptors in nearby body regions along the length of the body and carry motor signals from the spinal cord back to those regions (Fig. 35.17).
35.4.2 Nervous systems have voluntary and involuntary components.
As bodies with distinct internal organ systems evolved, two separate components of the nervous system emerged. When a gazelle senses a predator, some nerve circuits send a signal to run, an action that is under conscious control, while others signal the heart to beat faster and blood vessels supplying muscles to dilate, actions that occur unconsciously. Conscious reactions are under the control of the voluntary component of the nervous system, and unconscious ones are under the control of the involuntary component. Voluntary components mainly handle sensing and responding to external stimuli, whereas involuntary components typically regulate internal bodily functions. Both nervous system components are found in invertebrate and vertebrate animals.
Let’s look at these components in insects and crustaceans. In these animals, a nerve circuit regulates foregut function together with nerve circuits that regulate other parts of the animal’s digestive system. These circuits are an involuntary component of an animal’s nervous system. Sensory structures such as the antennae and eyes transmit information to the animal’s brain, which allows voluntary responses to stimuli received by these sensory organs. Other invertebrates such as mollusks also have involuntary components that regulate the function of internal organ systems that are distinct from their voluntary sensory–motor system.
In vertebrates, the peripheral nervous system is divided into somatic (voluntary) and autonomic (involuntary or visceral) components. The somatic nervous system is made up of sensory neurons that respond to external stimuli and motor neurons that synapse with voluntary muscles. This system is considered voluntary because it is under conscious control. However, many reflexes are controlled at lower spinal levels (discussed in the following section) or by the brainstem, independent of conscious control by the central nervous system.
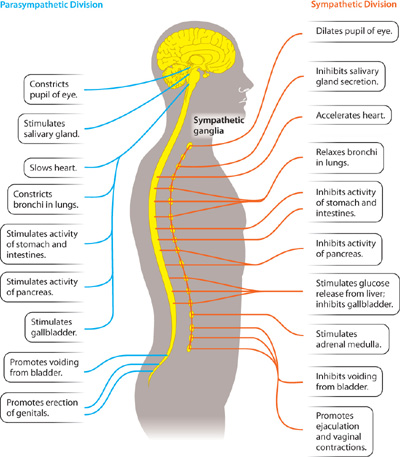
The autonomic nervous system (Fig. 35.18) controls internal functions of the body such as heart rate, blood flow, digestion, excretion, and temperature. It includes both sensory and motor components, which usually act without our conscious awareness. The autonomic nervous system, in turn, is divided into two major subdivisions, a sympathetic division and a parasympathetic division. Both divisions continuously monitor and regulate internal functions of the body. Generally, sympathetic and parasympathetic nerves have opposite effects, but not always. For example, sympathetic neurons stimulate the heart to beat faster, whereas parasympathetic neurons cause the heart to beat slower.
35-17
The sympathetic nervous system generally results in arousal and increased activity. It is the pathway activated when animals are exposed to threatening conditions, resulting in what is often referred to as the fight-or-flight response. This response includes an increase in the heart and breathing rates, increase in glucose release by the liver, and inhibition of digestion. The parasympathetic nervous system, in contrast, slows the heart and stimulates digestion as well as metabolic processes that store energy—in other words, it enables the body to “rest and digest.”
The sympathetic and the parasympathetic nerves have different anatomical distributions (Fig. 35.18) as well as different functions. Sympathetic nerves leave the CNS from the middle region of the spinal cord, forming ganglia along much of the length (thoracic and lumbar) of the spinal cord. Parasympathetic nerves leave from the brain by cranial nerves, as well as from lower levels (sacral) of the spinal cord.
35.4.3 The nervous system helps to maintain homeostasis.
The autonomic nervous system plays a key role in homeostasis, which is the ability to maintain a steady physiological state in the face of changing environmental conditions. Many physiological parameters are maintained in a narrow range of conditions, including pH, temperature, and ion concentrations. Homeostasis is maintained within individual cells as well as within the organism as a whole. Enzymes within a cell, for example, often work effectively only in narrow pH and temperature ranges. Ion concentrations also must be maintained within narrow ranges for normal cell function. As we have seen, the firing of action potentials by neurons requires particular ion concentrations on either side of the membrane. Similarly, the water content of cells and the body as a whole is kept stable through the careful regulation of ions and other solutes, as discussed in Chapter 41.
The concept of homeostasis was first described as regulation of the body’s “interior milieu” in the late 1800s by the French physiologist Claude Bernard, who is often credited with bringing the scientific method to the field of medicine. The term “homeostasis” was coined by the American physiologist Walter Cannon, whose book The Wisdom of the Body (first published in 1932) popularized the concept.
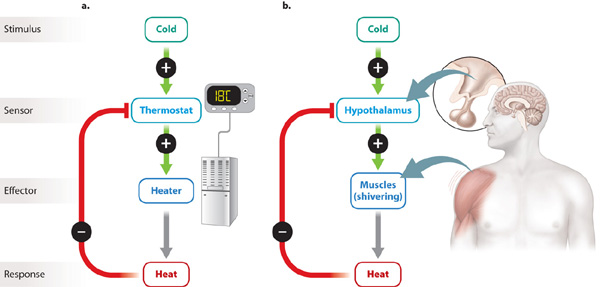
Maintaining steady and stable conditions takes work in the face of changing environmental conditions. That is, a cell (or organism) actively maintains homeostasis. How does the body, and in particular the nervous system, maintain homeostasis? Homeostatic regulation often depends on negative feedback (Fig. 35.19). In negative feedback, a stimulus acts on a sensor that communicates with an effector, which produces a response that opposes the initial stimulus. For example, negative feedback is used to maintain a constant temperature in a house. Cool temperature (the stimulus) is detected by a thermostat (the sensor). The thermostat sends a signal to the heater (the effector), producing heat (the response). In this way, a stable temperature is maintained (Fig. 35.19a).
35-18
In a similar fashion, humans and other mammals maintain a steady body temperature even as the temperature outside fluctuates. Nerve cells in the hypothalamus (located in the base of the brain) act as the body’s thermostat (Fig. 35.19b). A drop in body temperature signals the hypothalamus to activate the somatic nervous system to induce shivering and metabolic heat, as discussed in Chapter 40. At the same time, the hypothalamus activates the autonomic nervous system, causing peripheral blood vessels to constrict. The reduction in blood flow near the body’s surface in turn reduces heat loss to the surrounding air. An increase in temperature signals sweat glands and vasodilation of peripheral blood vessels to aid heat loss from the skin.
Homeostatic regulation, therefore, relies on negative feedback to maintain a set point, which in this case represents an animal’s preferred body temperature. The ability to maintain a constant body temperature is known as thermoregulation, and it is just one of many parameters that the body actively maintains, as we discuss in subsequent chapters.
35.4.4 Simple reflex circuits provide rapid responses to stimuli.
An animal that has perceived a predator has an advantage if it can move more quickly than the predator. Fast responses are made possible by simple reflex circuits that bypass the brain by directly connecting sensory neurons (detecting the presence of the predator) with motor neurons (the quick movement necessary to escape the predator). Reflex circuits are common in both the somatic and autonomic components of the nervous system.
Simple spinal reflex circuits connect sensory and motor neurons directly in the spinal cord so that a vertebrate can respond rapidly to sensory stimuli without the need for delay-causing conscious input from the brain. The large Mauthner neurons of fish are an example of such a circuit. Sensory neurons respond to threatening visual, tactile, or vibratory cues from the environment and transmit signals to the hindbrain of a fish, activating a large Mauthner neuron on the side opposite the stimulus. The Mauthner cell in turn rapidly activates motor neurons along the body, causing the fish to bend away from the threatening stimulus to initiate a rapid escape. At the same time, the Mauthner cell and motor nerves on the other side of the fish are inhibited. The giant axons of squid are part of a similar reflex circuit that allows them to swim quickly away from a predator.
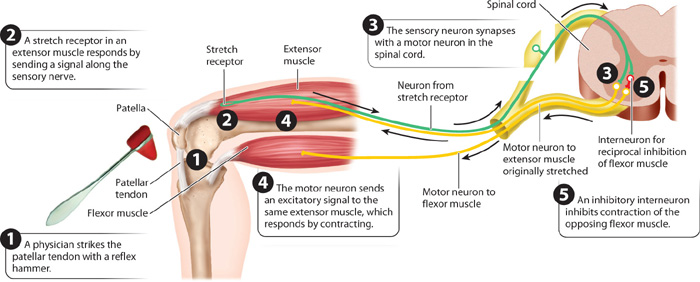
The knee-jerk reflex in humans is an example of a simple nerve circuit that includes only a single synapse between two neurons—a sensory neuron and a motor neuron (Fig. 35.20). Physicians commonly use this reflex to evaluate peripheral nervous and muscular system function. Let’s follow this reflex pathway. It starts with specialized “stretch” receptors in the extensor muscles of the leg, including the quadriceps muscle. These stretch receptors sense the stretch of the muscle that occurs during movement or in response to a physician’s strike of a reflex hammer on the stretch receptors. The receptors are part of a sensory nerve with dendrites that extend from the stretch receptors to cell bodies in ganglia beside the spinal cord. These cell bodies have axons that extend into the spinal cord. In response to a stretch, a signal is sent from the stretch receptor, through dendrite and cell body, to the axon. In the spinal cord, the axon of the sensory neuron synapses directly on motor neurons that travel from the spinal cord back to the muscle where the stretch originated. The signal from the muscle stretch receptor stimulates the motor neurons to increase the activation of the muscle, causing the muscle to contract and the leg to extend at the knee. This motion is the knee-jerk response.
35-19
This reflex arc does not include an interneuron: It is composed of just two neurons and one synapse. It provides a rapid change in muscle contraction because only one synapse is required to relay the sensory information back to the muscle. Neural circuits containing more neurons and synapses take longer to transmit nerve signals because transmission is delayed by communication at the synapses.
As well as being a useful test in examination, the knee-jerk reflex has a normal physiological role. In running or landing from a jump, the knee joint is flexed (that is, bent), stretching the quadriceps muscles that extend (that is, straighten) the knee joint. Knee flexion allows rapid adjustments in muscle force and knee position that help stabilize the body during running and jumping.
Because muscles can only contract, opposing movements at a joint such as flexion and extension require flexor and extensor muscles on either side of the joint (Chapter 37). These are often activated out of phase (that is, at different times) so that, when one set of muscles is activated (contracting), the other is inhibited (relaxed). The alternating movements of the limbs during walking and running provide an example. This pattern of joint and limb movement is achieved by reciprocal inhibition: When stretch receptors of the knee extensor muscles are activated to stimulate these muscles to extend the knee, they also inhibit the activity of opposing muscles that flex the knee (Fig. 35.20).
Reciprocal inhibition of opposing sets of muscles occurs in the spinal cord. Axons of the stretch receptor neurons not only synapse with motor neurons of extensor muscles but also synapse with inhibitory interneurons that inhibit motor neuron stimulation of the opposing flexor muscles. This inhibitory reflex pathway contains two synapses, one between the sensory neuron and interneuron and the second between the interneuron and the motor neuron to the flexor muscle.
Reciprocal inhibition also operates between the right and left sides of the body. For example, reciprocal inhibition helps to control the movement of the right and left limbs. In this case, interneurons cross the spinal cord to control the timing of activity by flexor and extensor muscles of the opposite limb. Reciprocal inhibition is also involved in the Mauthner cell circuit of fish to ensure that only one side of the fish’s body bends away from the stimulus to escape. The local spinal circuits that provide reciprocal inhibition are therefore fundamental to the alternating motion of the body and limbs that characterizes the movements of most animals.
In other cases, more complex circuits in the brain may act to coordinate sensory input and motor output. These circuits integrate other sources of sensory information, such as vision and balance, with conscious commands from the brain to voluntarily control motor behavior. In the next chapter, we look more closely at these sensory circuits.
35-20