37.4 VERTEBRATE SKELETONS
Although both vertebrate skeletons and invertebrate exoskeletons are rigid and have flexible joints, certain features distinguish them. Here, we explore the biology of vertebrate skeletons, focusing on how bones develop and grow and how they are repaired and remodeled.
37.4.1 Vertebrate bones form by intramembranous and endochondral ossification.
Vertebrate bones form and develop by two distinct processes. Bones of the skull and the ribs form when precursor cells differentiate into osteoblasts that immediately begin producing bone as a skeletal sheet or membrane. These bones are often referred to as membranous bones.
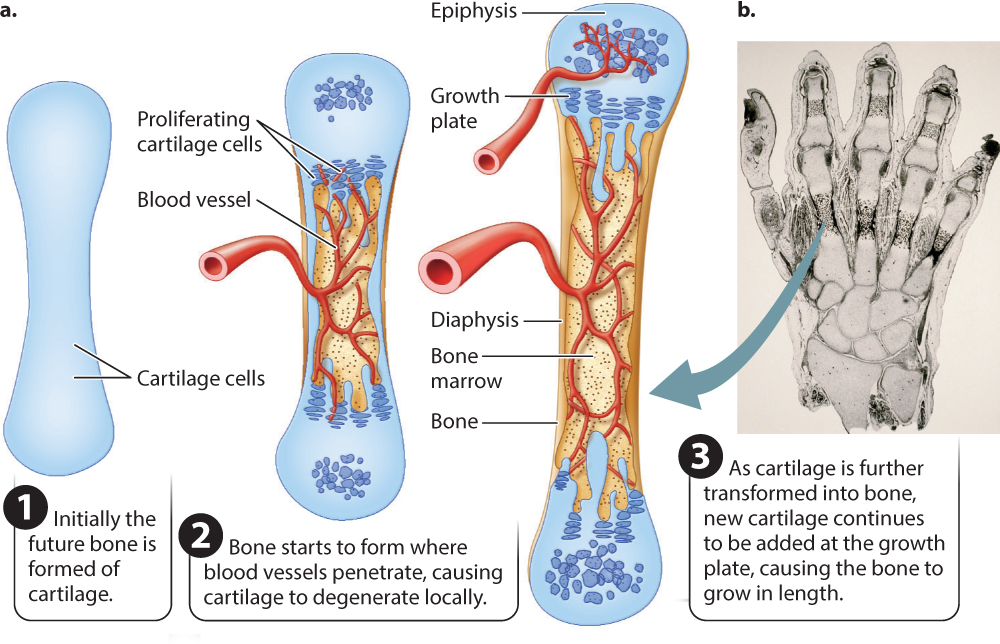
Most other bones of the body—the vertebrae, shoulder, pelvis, and limb bones—are formed as cartilage first (Fig. 37.18). The precursor cells initially become cartilage-producing cells, known as chondroblasts, that in the embryo grow into cartilage models of most bones of the developing body (Fig. 37.18a). For example, at approximately 9 weeks’ development, a human fetal hand consists mainly of this cartilage model (Fig. 37.18b). This embryonic cartilage is the same as the articular cartilage that remains at the ends of bones to form the joint surfaces in the growing and mature animal. Because cartilage is a pliable fluid-filled matrix, it grows by expansion within the structure as well as at the surface, enabling the fetal skeleton to grow rapidly. As the fetus grows and its skeleton matures, the cartilage model of the bone is invaded by blood vessels. The presence of blood vessels triggers the transformation of cartilage into hard, mineralized bone.
37-17
The cartilage template of a bone is replaced by one of two main types of bone tissue (Fig. 37.19). Both types contribute to the overall form of many bones in the body. Compact bone forms the walls of the bone’s shaft. It consists mainly of dense mineralized bone tissue containing bone cells known as osteocytes and the network of blood vessels supplying them with oxygen and nutrients.
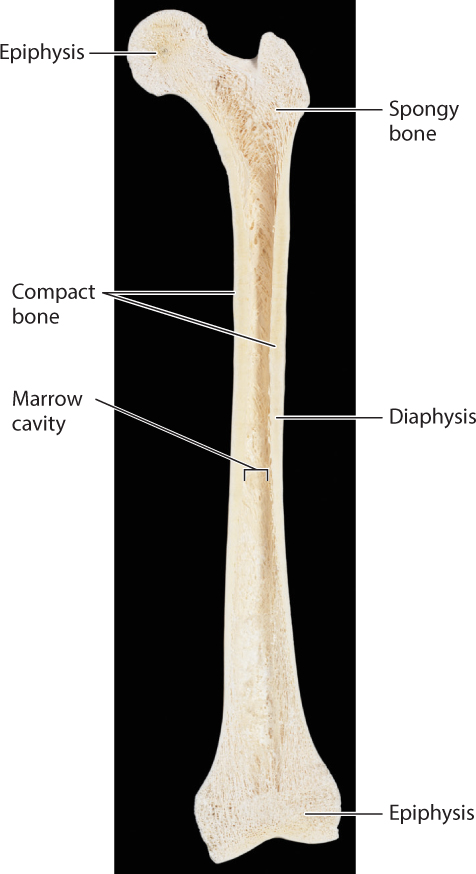
A second type of bone, called spongy bone, consists of small plates and rods known as trabeculae with spaces between them. Spongy bone is found in the ends of limb bones and within the vertebrae. It reduces the weight of bone in these regions, but it is also stiff so that forces are transmitted effectively from the joint to the bone shaft. Bone marrow, a fatty tissue found between trabeculae and also within the bone’s central cavity, contains many important cell populations, including blood-forming cells, other stem cells (Chapter 20), and immune system cells (Chapter 43), as well as fat cells.
Bones can grow in length as well as in width. Growth in length occurs at a growth plate (see Fig. 37.18), a region of cartilage between a middle region, called the diaphysis, and the end, called the epiphysis. Blood vessels invade the central diaphysis region, as well as each epiphysis, triggering the transformation of cartilage into bone. The growth plate is left in between. The growth plate adds new cartilage toward the bone’s diaphysis, enabling bone length to continue to increase after birth (see Fig. 37.18). At maturity, the growth plates fuse as the remaining cartilage is replaced with bone, preventing further growth in length. The fusion of growth plates is typical of most mammals and birds. In amphibians and reptiles, the growth plates often do not fuse and growth may continue at a slower pace over much of their lifetime.
Limb bones grow in diameter when osteoblasts deposit new bone on the bone’s external surface. At the same time, bone is removed from the inner surface, expanding the marrow cavity. Bone removal is slower than bone growth, thickening the walls during growth. Bone is removed from the marrow cavity by a group of cells called osteoclasts that secrete digestive enzymes and acid to dissolve the calcium mineral and collagen. These dissolved compounds are reabsorbed and recycled for bone formation in other regions of the skeleton. Through this process, the vertebrate skeleton serves as an important store of calcium and phosphate ions. For example, in female birds, calcium removed from the skeleton is regularly used to form the eggshell.
A great advantage of endoskeletons compared with exoskeletons is that a vertebrate’s skeleton can be repaired if damaged by osteoblasts and osteoclasts forming and removing mineralized tissue in particular regions of the bone. Generally, physically active younger adults have thicker bones than less-active individuals. However, in older humans, bone tissue is gradually lost as osteoclasts remove more bone than is produced by osteoblasts. Bone loss is particularly severe in women after menopause, in part because of hormone shifts, and can lead to osteoporosis, a condition that significantly increases the risk of bone fracture.
37.4.2 Joint shape determines range of motion and skeletal muscle organization.
The shapes of the bone surfaces that meet at a joint determine the range of motion at that joint. Joints range from simple hinge joints that allow one axis of rotation to ball-and-socket joints that allow rotation in three axes (Fig. 37.20). The human elbow joint and the ankle joint of a dog are examples of hinge joints. The shoulder and hip joints are examples of ball-and-socket joints, which allow the widest range of motion, as when you throw or kick a ball. The joints at the base of each finger are intermediate: They allow you to flex and extend your fingers as well as spread them laterally or move them together when making a fist or grasping different-sized objects.
37-18
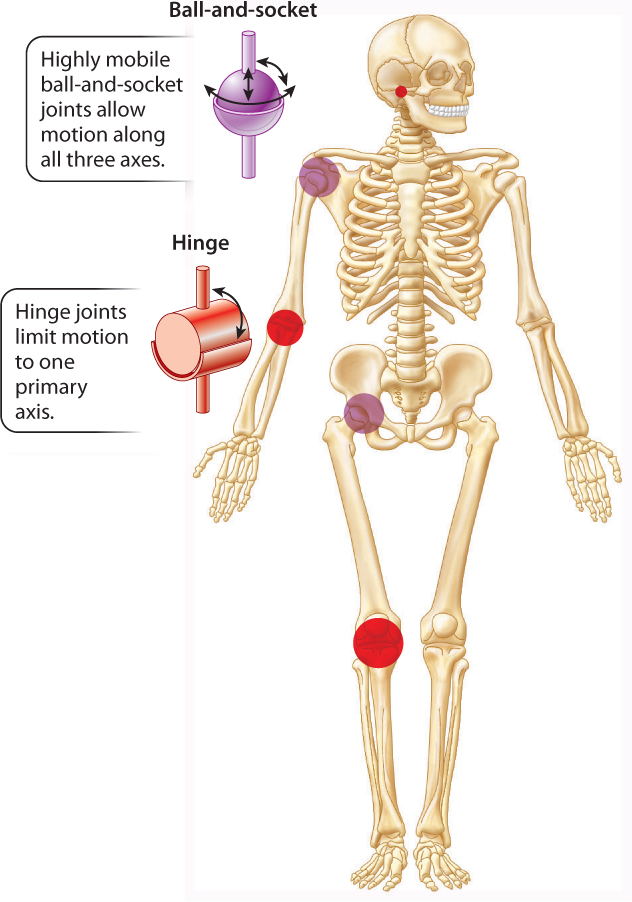
Joints with a broader range of motion are generally less stable. The shoulder is the most mobile joint in the human body, but it is also the most often dislocated or injured. In contrast, the ankle joint of dogs, horses, and other animals is a stable hinge joint that is unlikely to be dislocated, but its range of motion is limited to flexion and extension. Because muscles are arranged as paired sets of antagonists to move a joint in opposing directions, hinge joints are controlled by as few as two antagonist muscles (generally referred to as a flexor and an extensor). In contrast, ball-and-socket joints minimally have three sets of muscle antagonists to control motion in three different planes. As a result, a more complex organization of muscles is needed to control the movements of the arm at the shoulder joint or the leg at the hip joint.
37.4.3 Muscles exert forces by skeletal levers to produce joint motion.
By serving as a rigid set of levers, the skeleton enables muscles to transmit forces that cause joint rotation. Analogous to the gears of a bicycle, muscles that attach farther from a joint’s axis of rotation produce slower but stronger movements, whereas muscles that attach closer to a joint produce rapid but weaker movements. This is why you open a door by pulling on a doorknob located opposite from its hinges. The doorknob’s location increases the lever distance, making it easier to pull the door open than if it were placed next to the hinge.
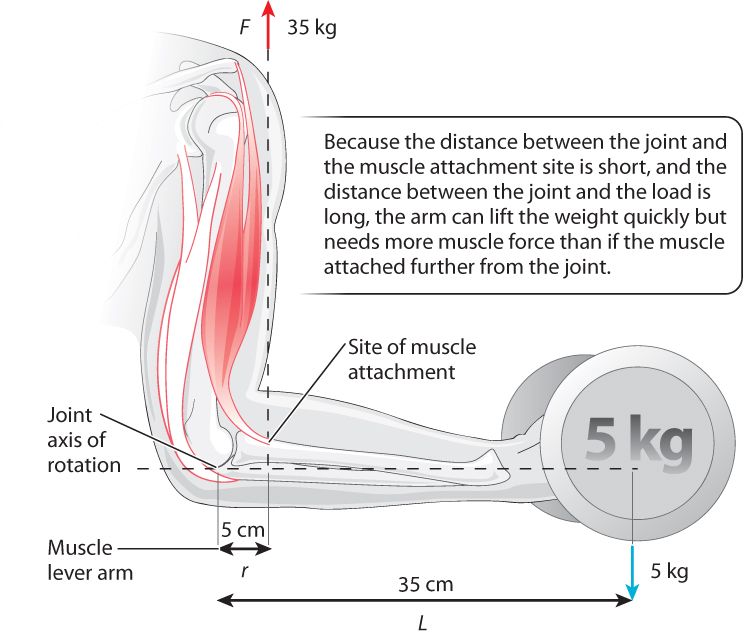
The strength of rotational movement, or torque, is determined by the product of a muscle’s force (F) and perpendicular lever distance (r) from the axis of joint rotation: F x r. Muscles produce joint torques to counteract torques that are applied by external forces, such as a weight that is carried by your hand. In Fig. 37.21, the axis of rotation is the elbow, and contraction of the biceps muscle causes the arm to flex at the elbow. The strength of elbow rotation is determined by the biceps muscle’s force (F) and the lever distance between the elbow joint and the site of muscle attachment, where the muscle pulls on the arm bone (r). Muscles produce stronger rotational movements either by exerting a larger force or by having a longer lever distance between the joint and muscle attachment site. Muscles that transmit force closer to the joint (smaller lever distance) result in less rotational strength but can produce large ranges of movement compared to muscles that attach farther from the joint.
37-19
As a result, the trade-off of force versus velocity that we observed earlier as a basic contractile property of a muscle also applies to the way muscles pull on the skeleton to cause joint movement. Muscles that transmit large forces for body movement and weight support, such as the calf muscles that extend your ankle joint, typically have large leverage for strong joint rotation. The heel of your foot gives the Achilles tendon of your calf muscles this greater leverage.
Animal joints located a long distance from the muscle attachment site (high muscle leverage) and a shorter distance from where the output force is exerted produce strong but slow movements. Moles, armadillos, and spade-foot toads are examples of digging animals that have specialized muscle–joint arrangements like this (Fig. 37.22). In contrast, joints located a short distance from the muscle attachment site (low muscle leverage) and a longer distance from where the output force is exerted produce faster movements but with less force. These are found in animals adapted for high-speed running, such as antelope, cheetahs, and horses.
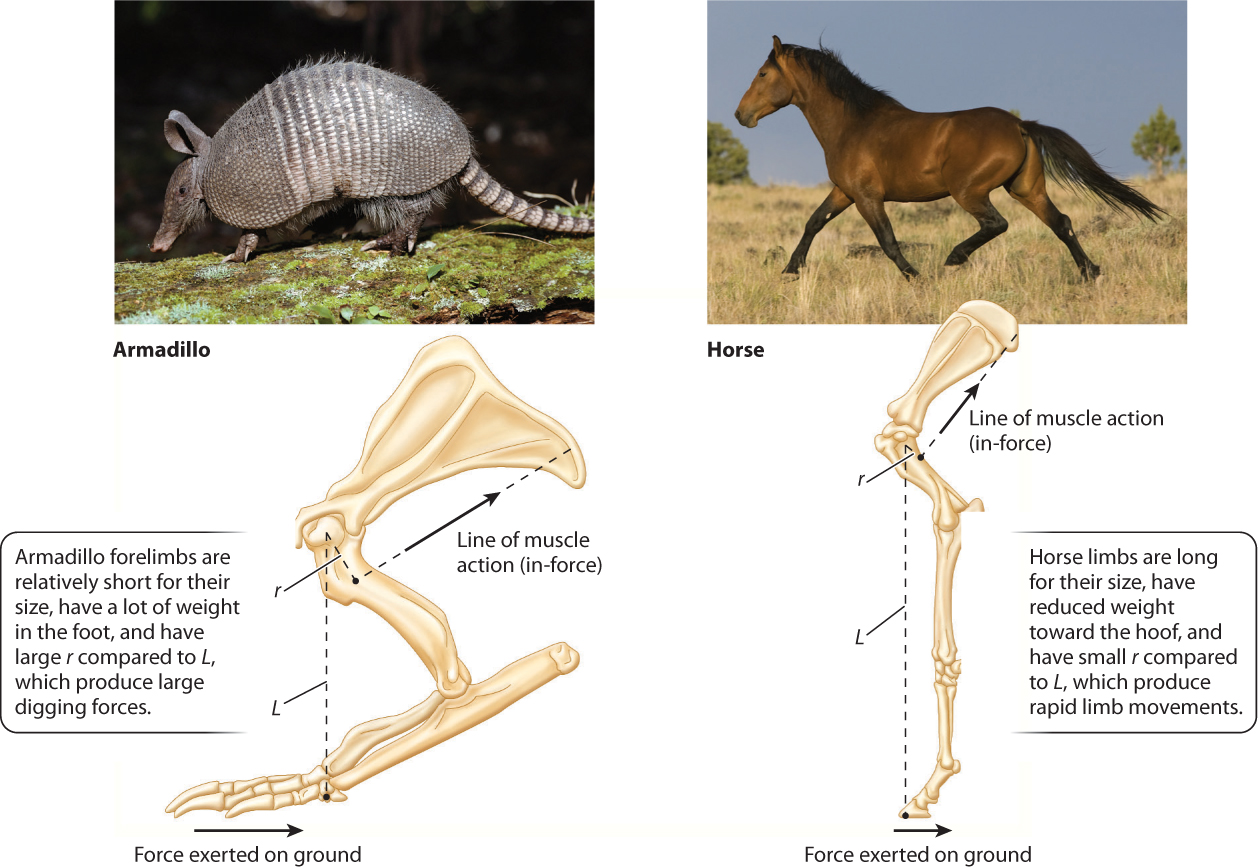
Larger, faster animals also increase their economy of movement by reducing the mass of muscles and bones toward the end of their limbs. This lowers the energy cost of swinging the limb, which antelope rely on for fast economical locomotion to escape predators. Predators generally have heavier limbs that require larger muscles to move but which make them stronger.
37-20